Background information about Chloroplast Organelle DNA
Plastids exist in plants and algae. They start as undifferentiated proplastids and may develop into:
- Amyloplasts: (starch storage)
- Chloroplasts: (photosynthesis)
- Etioplasts: (chloroplasts not exposed to light)
- Elaioplasts: (storing fat)
- Chromoplasts: (pigment synthesis and storage)
- Leucoplasts/Leikoplasts: (monoterpene synthesis)
In algae, the term leucoplast (leukoplast) is used for all unpigmented plastids. Etioplasts, amyloplasts and chromoplasts do not occur in algae. Plastids are similar to mitochondria in that both contain circular DNA (plastome). Chloroplast DNA is abbreviated as ctDNA, sometimes as cpDNAa or sometimes as plastid DNA or ptDNAb (mitochondrial DNA is usually abbreviated as mtDNA). It should be noted that plasmids most often also have a circular DNA form. As sperm is small and contains mostly DNA, while the ovum is large and contains much cytoplasm containing both mitochondria or chloroplasts, both the chloroplasts and the mitochondria are maternally inherited. mtDNA contains about 20 genes.
ctDNA contain about 100 genes to synthtsize proteins:
- DNA polymerases
- RNA polymerases
- rRNA
- ribosomal proteins
- tRNA
- aminoacyl-tRNA-synthetases
- enzymes to synthesise chlorophyl and carotenoids
- ORFs (Open Reading Frames) with start and stop codons to encode proteins, many for photosynthesis)
Introns are included in the nucleotide sequences of plastids.
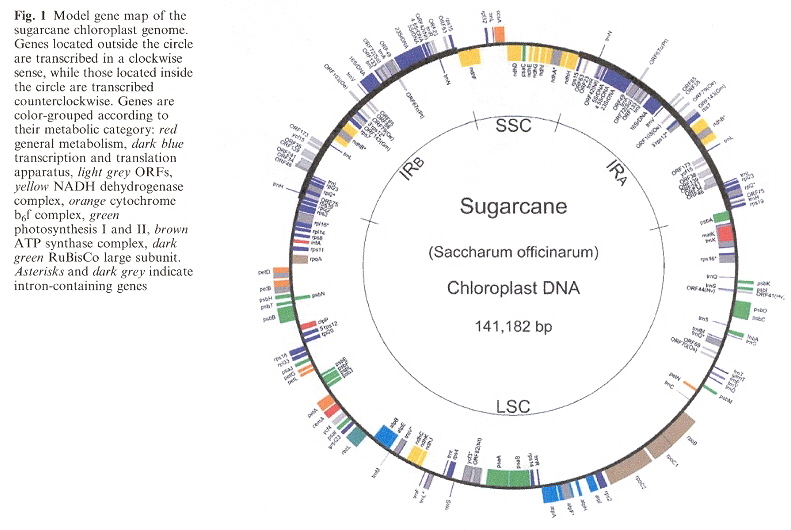
Example of Sugarcane Plastomec
According to the endosymbiotic theory, eukaryotic cells incorporated prokaryotic photosynthetic eubacteria (most likely cyanobacteria) by invagination, that became plastids (similarly, α -protobacteria were incorporated as mitochondria before chloroplasts were incorporated). In fact, the endosymbiont in cryptomonads and chlorarachniophyte algae retains vestigial cytoplasm and a nucleus. Both mitochondria and chloroplasts specialized in functions of energy production as symbionts (for example, CO2 plus light, results in sucrose in Chloroplasts, while mitochondria use glucose to produce ATP). Genetic information (DNA) then transferred between mitochondria, chloroplasts, and the nucleus. Endosymbiotic development resulted in chloroplast encoded nuclear proteins targeting chloroplast stroma, thylakoid membranes, etc. DNA incorporated into nuclear DNA from plastids became highly fragmented, and over time much of this DNA is eliminated from the nucleus. There is a lack of agreement as to how many endosymbiotic events took place (chloroplasts and mitochondria implying one or two events, or more), criteria being proposed ("Organelle Genes and Genomes", by N. W. Gillham, Oxford U.P., 1994, p. 37):
- Evolutionary histories of nuclear and organelle genomes are different and known with certainty
- Evolutionary history of nuclear genome differs from at least one organelle, although history of nuclear genome history is not certain
- If it could be shown that mitochondria and chloroplasts arose from different lineages of bacteria
- Chloroplasts and mitochondria are of polyphyletic origin
Not only does genetic information move between the organelles and the nucleus (both directions between nucleus and mitochondria), but it also transfers horizontally: ( interspeciesd transmission). This is often referred to as "promiscuous" DNA transfer. mtDNA incorporated into the nuclear DNA is named numtDNA, while ctDNA incorporated into the nuclear DNA is named nuptDNA (sometimes, nupDNA)b.
Evolution of Organelles: DNA Passes between Nucleus and other Organelles
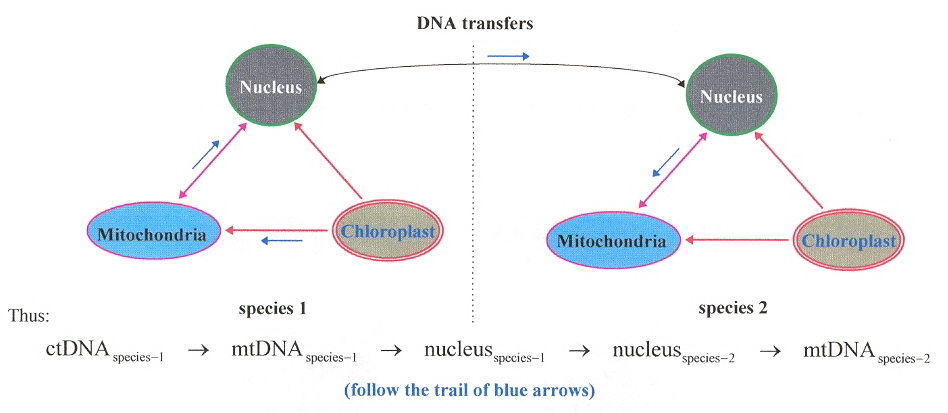
numtDNAb
Evolution of numtDNAb
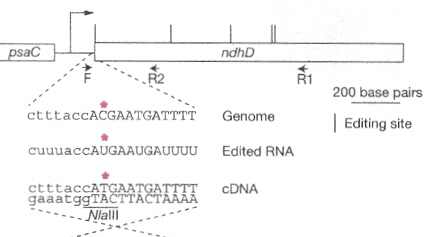
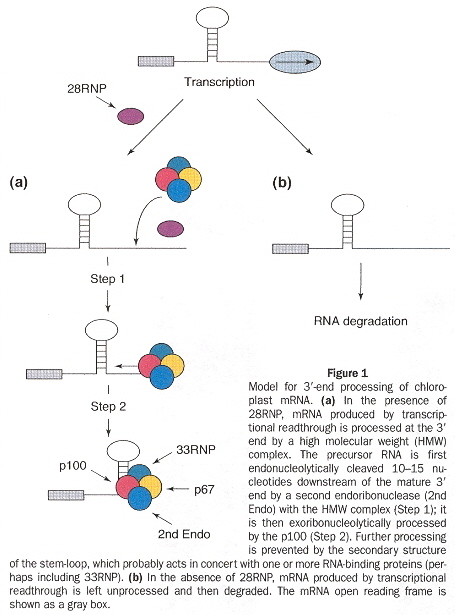
Enzymes originating from the nuclear DNA, in the cytosol can pass the chloroplast membranes. An example of the close coordination required between nuclear and cytoplasmic genetic components, consider that chloroplast mRNAs require nuclear encoded factors (as well as light) to activate mRNA translation. This has been illuminated by mutations affecting the secondary structure of chloroplast RNAe (also called cpRNAf). Note that there are special binding sites such as RBS (ribosome-binding sites). Another example of RNA editing is the insertion, deletion, or modification of nucleotides. As an example, the conversion of cytidine into uridine (often resulting in the restoration of conserved amino acidsg).
Translational Expression of RNA from ctDNA
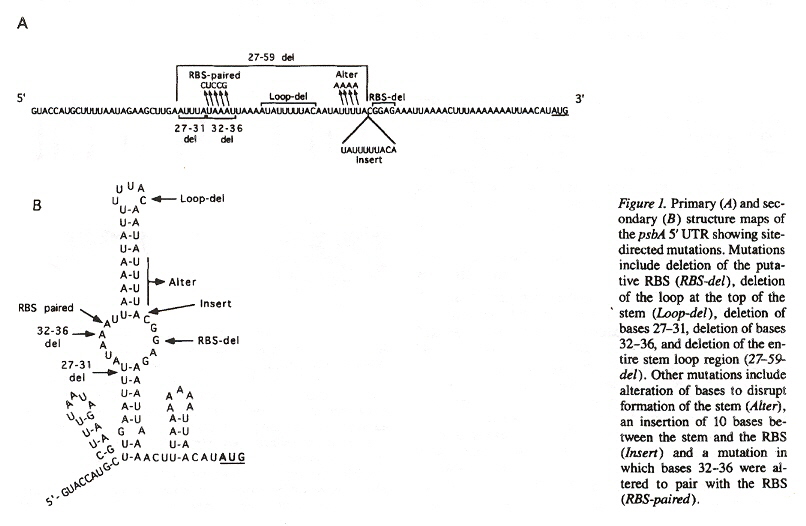
RNA Editing
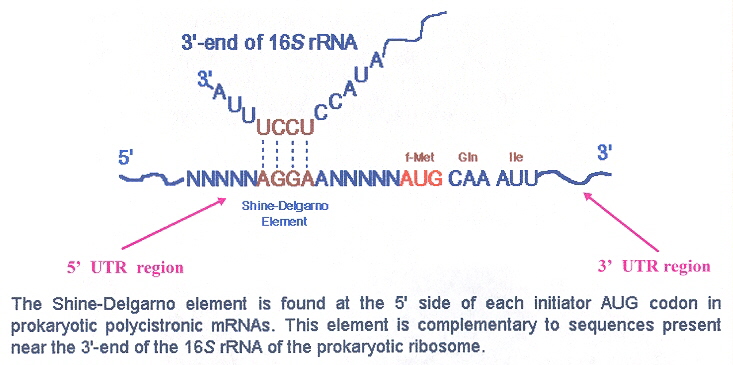
The initiation codon for translation of plastid mRNA is AUG (although there are exceptions, such as GUG, UUG, ATT, ATA, etc.). Ribosomal rRNA can interact directly with translation initiation through base-pairing at the Shine-Delgarno sequence circa 10 to 30 nucleotides towards the 5' end of the plastid mRNA. Deletion of the Shine-Delgarno sequence from plastid mRNA can destroy translation (S-D sequences are not an absolute requirement for plastid mRNA translation). Other UTR (untranslated regions) mechanisms operate to allow plastid mRNA translation, but will not be discussed here, other than to note that 3' UTRs are important, not just 5' UTRs. Note that plastid mRNA have inverted repeat loops which fold into stem-loops at the 3' terminal. The function of the stem-loop structures is to prevent degradation. The stability conferred (most likely by RNA-binding proteins) increases the half-life from 8 to more than 40 hours (for psbA)h. These stem-loops act as a transcriptional terminator in bacteria, but do not act as transcriptional terminators in chloroplastsi. While the Shine-Delgarno sequence finds the translation initiation site, the terminal translation site is one of the usual nonsense stop codons, which allows a release factor to release the associated completed polypeptide. Mitochondrial translation works in the same fashion as that for plastids, but while in eubacterial systems translation is usually by repression, in organelles, translation is usually by activation (but of course, there are exceptions)j.
mRNA Translation Interaction at the Shine Delgarno Sequence
mRNA 3'-UTR Stem-Loop Stability
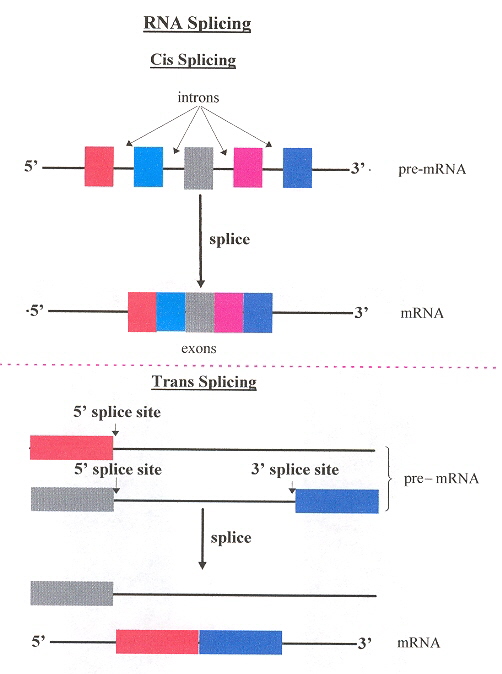
To clarify questions of terminology, two kinds of RNA splicing are commonly refered to. Cis splicing removes introns from a single pre-RNA strand. Trans splicing collects segments of pre-RNA but from different pre-RNA strands.
Cis/Trans Splicing
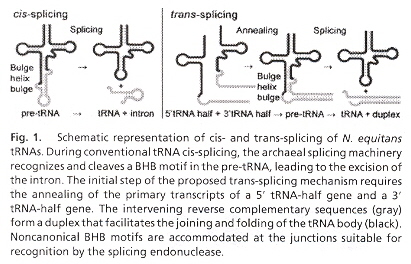
An interesting example is the splicing endonuclease of Nanoarchaeum equitans which acts to remove introns from intron-containing precursor tRNA (pre-tRNA). The result is two tRNA half-molecules: a 51 half-exon ending with a 21, 31 -cyclic phosphate, and a 31 half-exon with a 51 -hydroxyl terminus. Then both halves of the tRNA exons are ligated. The splicing endonuclease recognizes a consensus bulge-helix-bulge (BHB) structure found at intron-exon junctions. Both cis-splicing as well as trans-splicing mechanisms have been found k.
tRNA cis- and trans-splicing
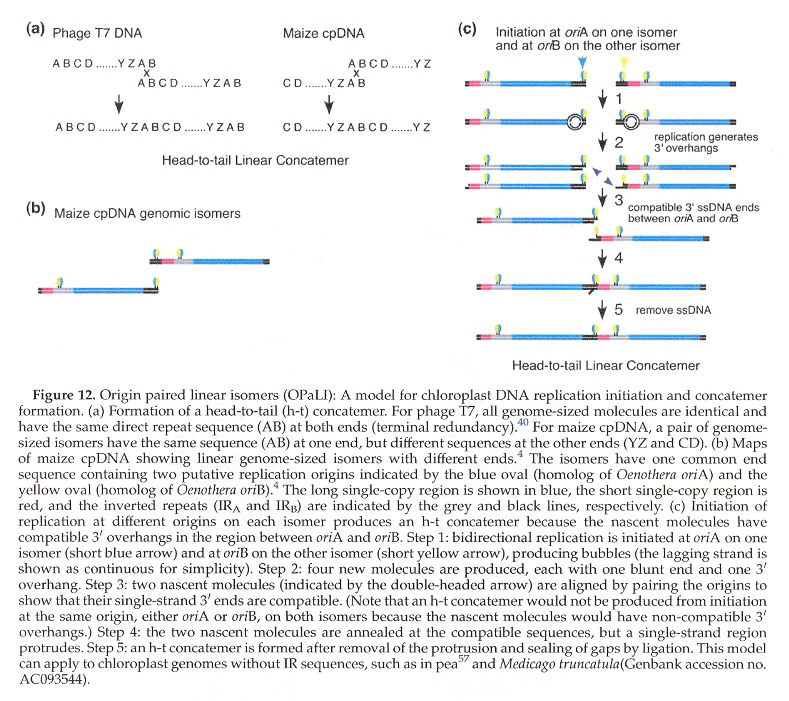
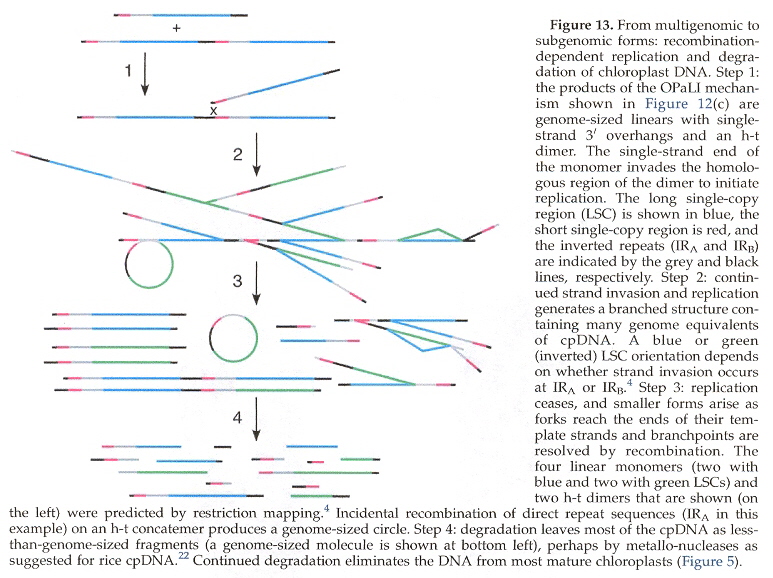
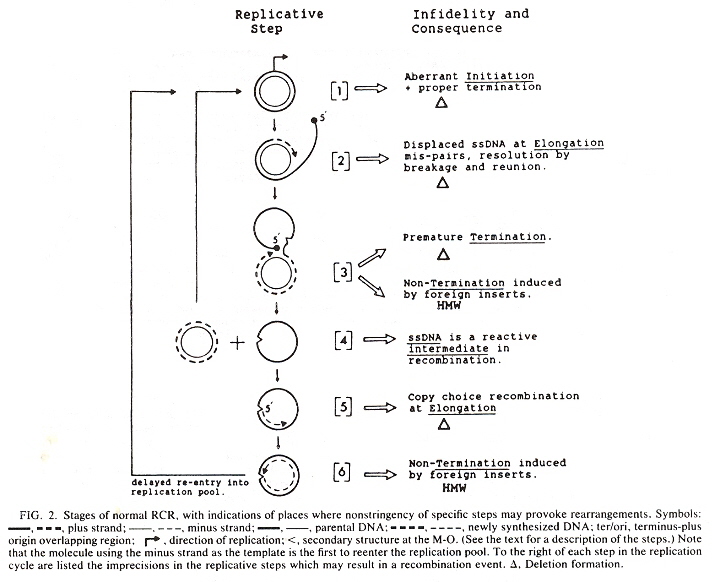
The standard model of cpDNA replication is rolling circle theta replication. This is discussed in the next listed item and should be reviewed. However, there are differing opinions about replication. Another possibility is head-to-tail linear concatemersl.
cpDNA Replication: Linear Concatemers (non-standard model)
cpDNA Replication Ceases
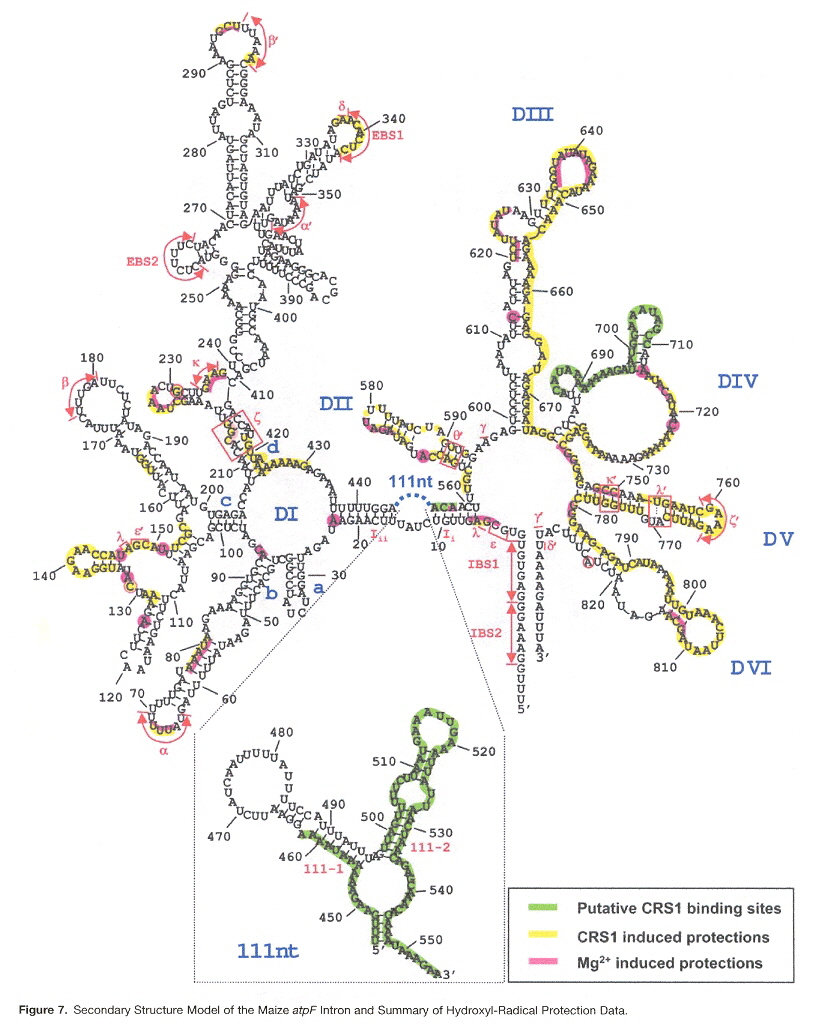
Many introns have lost the capacity to self-splice. As an example, some group II introns (catalytic RNAs found especially in algae and plants in chloroplasts, as well as some mitochondria) require nuclear-encoded splicing proteins. In particular, Zea Mays has CRS1 or Chloroplast RNA Splicing 1 protein required by the chloroplast intron on gene atpF. Group II introns are typically composed of six helical domains eminating from a central core. CRS1 has repeated domains called CRM domains (chloroplast RNA splicing and ribosome maturation binding domains). CRS1 binding at domains I and IV causes a shift in intron domain population: the average structure becomes more compact as expected of a catalytically active intron. This suggests that CRS1 aids in intron foldingm.
Maize atpF Intron
a "Chloroplast DNA variation of Quercus rubra L. in North America and comparison with other Fagaceae", by C. R. Magni, A. Ducousso, H. Caron, R. J. Petit, A. Kremer, Molecular Ecology, Feb. 2005, 14, 2, 513 - 524
b "Mutational Decay and Age of Chloroplast and Mitochondrial Genomes Transferred Recently to Angiosperm Nuclear Chromosomes", by C. Y. Huang, N. Grünheit, N. Ahmadinejad, J. N. Timmis, W. Martin, Plant Physiological Preview, June 10 2005, 138, 1723 - 1733
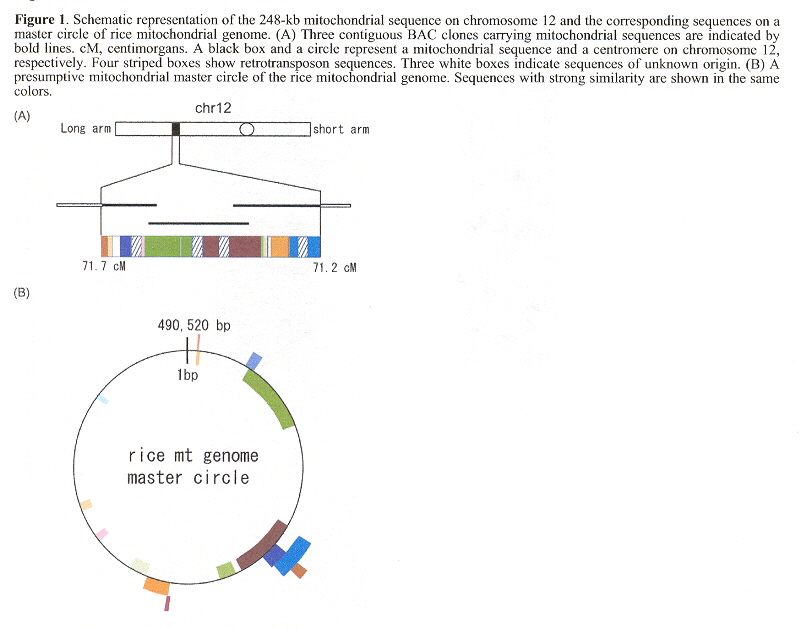
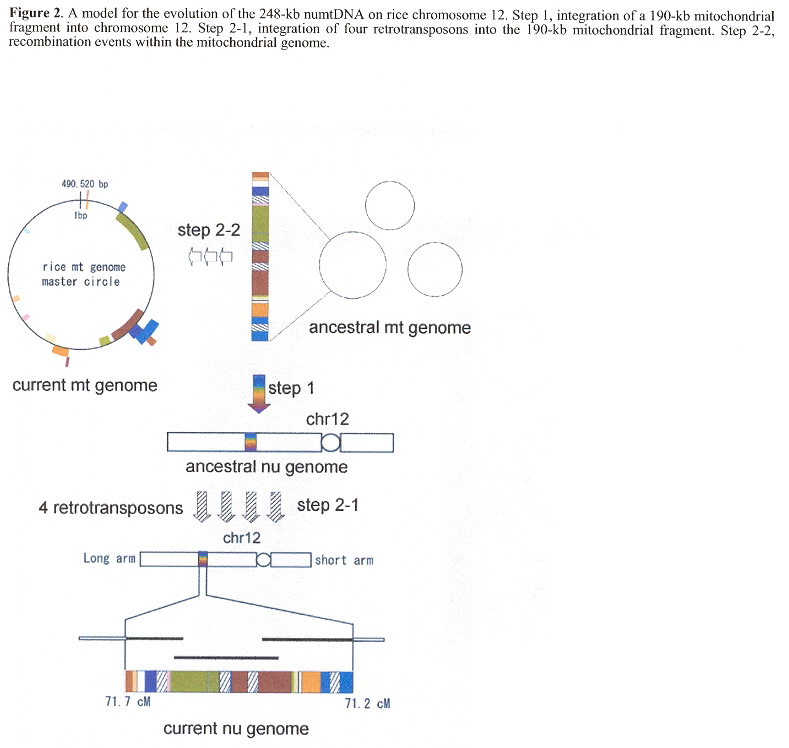
"Translocation of a 190-kb mitochondrial fragment into rice chromosome 12 followed by the integration of four retrotransposons", by M. Ueda, N. Tsutsumi, K-i. Kadowaki, International journal of Biological Sciences, 2005, 1, 3, 110 - 113
"The Rice Nuclear Genome Continuously Integrates, Shuffles, and Eliminates the Chloroplast Genome to Cause Chloroplast-Nuclear DNA Flux", by M. Matsuo, Y. Ito, R. Yamauchi, J. Obokata, The Plant Cell, March 2005, 17, 3, 665 - 675
c "Structural features and transcript-editing analysis of sugarcane (Saccharum officinarium L.) chloroplast genome", by T. C. Júnior, D. M. Carraro, M. R. Benatti, A. C. Barbosa, J. P. Kitajima, H. Carrer, Current Genetics, Dec. 2004, 46, 6, 366 - 373
This paper uses "plastome" in preference to ctDNA genome.
d "A fragment of chloroplast DNA was transferred horizontally, probably from non-eudicots, to mitochondrial genome of Phaseolus", by M. Woloszynska, T. Bocer, P. Mackiewicz, H. Janska, Plant Molecular Biology, Nov. 2 2004, 56, 5, 811 - 820
e "Translation of the psbA mRNA of Chlamydomonas reinhardtii Requires a Structured RNA Element Contained within the 5' Untranslated Region", by S. P. Mayfield, A. Cohen, A. Danon, C. B. Yohn, The Journal of Cell Biology, Dec. 1994, 127, 6, Part I, 1537 - 1545
f "Antisense Transcript and RNA Processing Alterations Suppressing Instability of Polyadenylated mRNA in Chlamydomonas Chloroplasts", by Y. Nishimura, E. A. Kikis, S. L. Zimmer, Y. Komine, D. B. Stern, The Plant Cell, Nov. 2004, 16, 11, 2849 - 2869
g "A pentatricopeptide repeat protein is essential for RNA editing in chloroplasts", by E. Kotera, M. Tasaka, T. Shikanal, Nature, Jan 20 2005, 433, 7023, 326 - 328
h "Degrading chloroplast mRNA: the role of polyadenation", by R. Hayes, J. Kudla, W. Gruissem, Trends in Biochemical Sciences, May 1999, 24, 5, 199 - 202
i "Polyadenylation and Degradation of mRNA in the Chloroplast", by G. Schuster, I. Lisitsky, P. Klaff, Plant Physiology, Aug. 1999, 120, 4, 937 - 944
j "Translation in chloroplasts", by W. Zerges, Biochimie, June-July 2000, 82, 6-7, 583 - 601
"Shine-Dalgarano-like sequences are not required for translation of chloroplast mRNAs in Chlamydomonas reinhardtii chloroplasts or in Escherichia coli", by D. C. Fargo, M. Zhang, N. W. Gillham, J. E. Boynton, Molecular and General Genetics, Feb. 1998, 257, 3, 271 - 282
"The Shine-Dalgarno-like sequence is a negative regulatory element for translation of tobacco chloroplast rps2 mRNA: an additional mechanism for translational control in chloroplasts", by W. Plader, M. Sugiuga, The Plant Journal, May 2003, 34, 3, 377 - 382
"Analysis of the Translational Initiation Region on the Euglena gracilis chloroplast Ribulose-bisphosphate Caurboxylase/Oxygenase (rbcL) Messenger RNA", by J. S. Koo, L. L. Spremulli, The Journal of Biological Chemistry, Mar. 11 1994, 269, 10, 7494 - 7500
k "The heteromeric Nanoarchaeum equitans splicing endonuclease cleaves noncanonical bulge–helix–bulge motifs of joined tRNA halves", by L. Randau, K. Calvin, M. Hall, J. Yuan, M. Podar, H. Li, D. Söll, Proceedings of the National Academy of Sciences U.S., Dec. 13 2005, 102, 50, 17934 - 17939
l "Changes in the Structure of DNA Molecules and the Amount of DNA Per Plastid During Chloroplast Development in Maize", y D. J. Oldenburg, A. J. Bendich, Journal of Molecular Biology, Dec. 10 2004, 344, 5, 1311 - 1330
m "CRS1, a Chloroplast Group II Intron Splicing Factor, Promotes Intron Folding through Specific Interactions with Two Intron Domains", by O. Ostersetzer, A. M. Cooke, K. P. Watkins, A. Barkan, The Plant Cell, Jam. 2005, 17, 1, 241 - 255
"Generation and evolutionary fate of insertions of organelle DNA in the nuclear genomes of flowering plants", by C. Noutsos, E. Richly, D. Leister, Genome Research, May 2005, 15, 5, 616 - 628
Organelle DNA from both mitochondria and plastids have been detected in the nuclear DNA of both Arabidopsis and rice. Detailed study of this DNA is summarized in this paper.
"Origin, evolution and genetic effects of nuclear insertions of organelle DNA", by D. Leister, TRENDS in Genetics, Dec. 2005, 21, 12, 655 - 663
In humans, some insertions of nuclear mitochondrial DNA into the nucleul DNA (nDNA) are associated with heritable diseases (Pallister-Hall syndrome, inherited factor (F)VII deficiency, MADH2 tumour suppressor mutation in colorectal carcinoma). The author notes that organelle DNA exists in different forms, such as mtDNA and ptDNA, and proposes a generic orgDNA. As organellar DNA gets inserted into the nucleus, such as numtDNA and nuptDNA, the author suggests a generic norgDNA. Apparently nucleus-to-chloroplast or mitochondrion-to-chloroplast transfer is extremely rare or never occurs. One mechanism by which orgDNA gets integrated into nDNA is during the repair of nuclear DNA double-stranded breaks. Other mechanisms include:
- organelle lysis during autophagy, gametogenisis or fertilization
- Fusion of mitochondria
- Nuclear inclusion of mitochondria or nuclear attachment of plastids
- Stromules a connecting plastids with mitochondria or the nucleus
a Stromules are projections that interconnect plastids that enable the transport of molecules.
Back to TopBackground information about Mitochondrial Organelle DNA
"Bidirectional Replication Initiates at Sites Throughout the Mitochondrial Genome of Birds", A. Reyes, M. Y. Yang, M. Bowmaker, I. J. Holt, Journal of Biological Chemistry, Feb. 4 2005, 280, 5, 3242 - 3250
Mitochondrial replication occurs in both directions around circular mtDNA with initiation sites flanking the major non-coding region (NCR). &theta replication consists of a single discrete replication origina and even if there exists multiple replication origins, it was assumed that replication initiation takes place at a discrete site for a given DNA region. It is expected that the vertibrate mtDNA replication mechanism is conserved. The strand-asynchronous model of mtDNA replication is based upon two strands originating from initiation sites, designated the heavy and the light strand origins (OH and OL) in which OL can form a hairpin stem-loop. However, something is wrong: avian mtDNA do not appear to have a hairpin stem-loop. Investigations indicate bidirectional replication forks at multiple sites traveling in both directions around the mtDNA circle.
Organization of Circular mtDNA
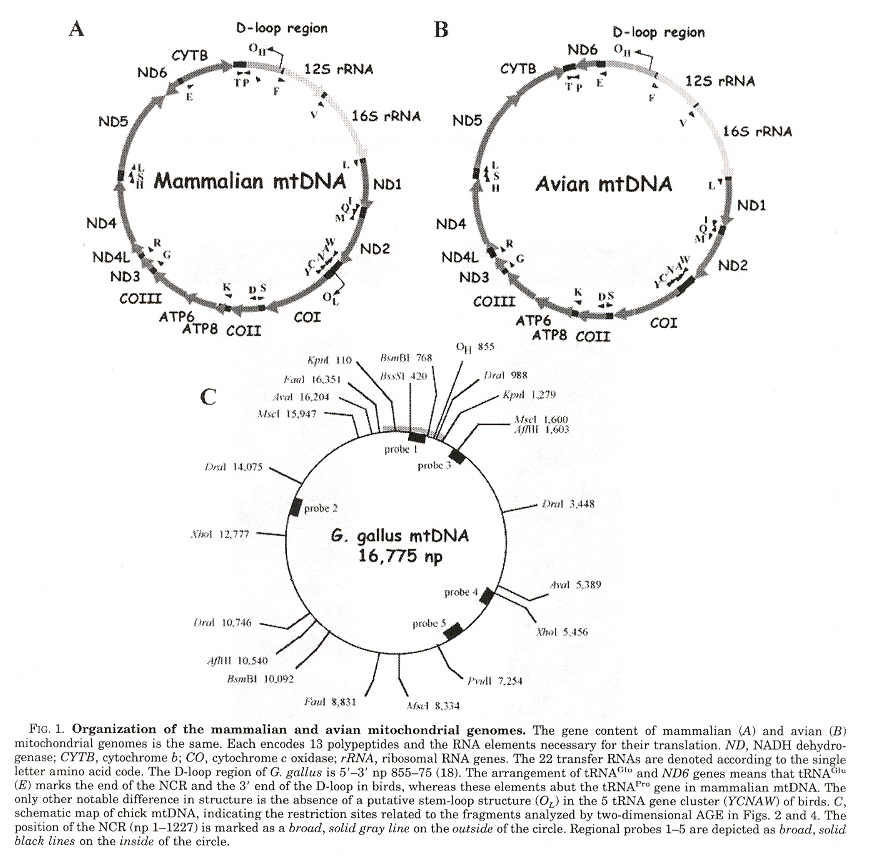
Figs. B, E, H show replication initiation at 3', Fig. K shows bidirectional initiation at 3' and 5'

Bidirectional initiation at multiple sites but not crossing the NCR or D-loop region
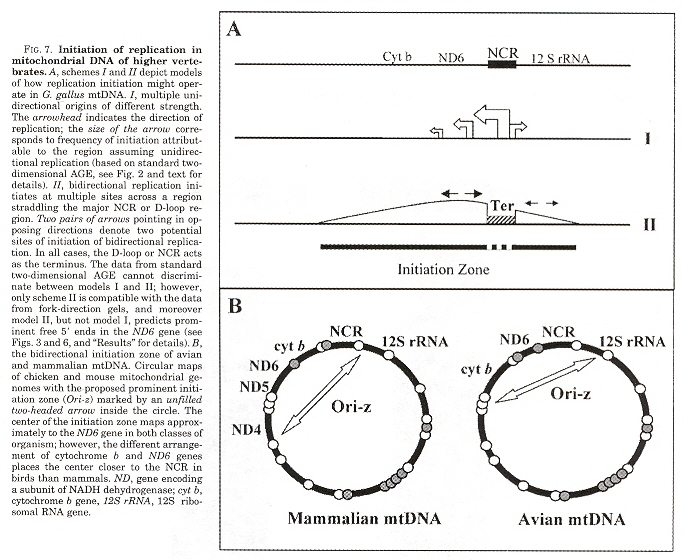
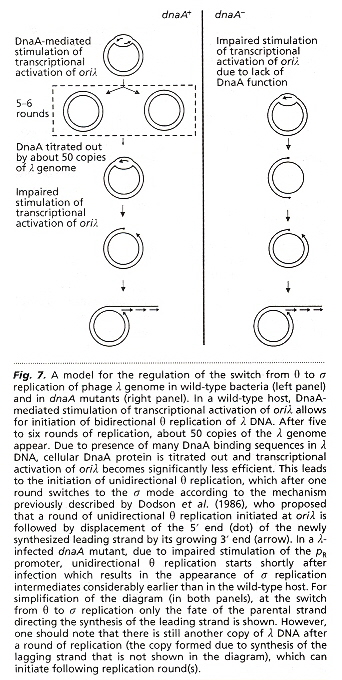
a "Regulation of the switch from early to late bacteriophage λ DNA replication", S. Barańska, M. Gabig, A. Węgrzyn, G. Konopa, A. Herman-Antosiewicz, P. Hernandez, J. B. Schvartzman, D. R. Helinski, G. Węgrzyn, Microbiology, 2001, 147, 535 - 547
θ replication proceeds by replicating circular copies of λ phage genome, using host DnaA protein, until there is insuficient protein, whereupon σ replication of λ phage genome is produced. The &lambda phage genome is linear dsDNA at 5' and 3' ends ligate to obtain the circular DNA. The linear σ phage dsDNA then concatenates multiple times.
Background information about Kinetoplast Organelle DNA
List of important kDNA morphological structures along with their abbreviations. It is significant that mammal mitochonria have been found to contain triple-stranded DNA ("Organelle Genes and Genomes", by N. W. Gillham, Oxford U.P., 1994, p. 58). This section is primarily about kDNA, but for the most part applies to mtDNA as well: mammal, angiosperm, insect, nematodes, cnidaria, algae, fungi, and sea anemone mtDNA not being that different than kDNA (ibid. p. 58).
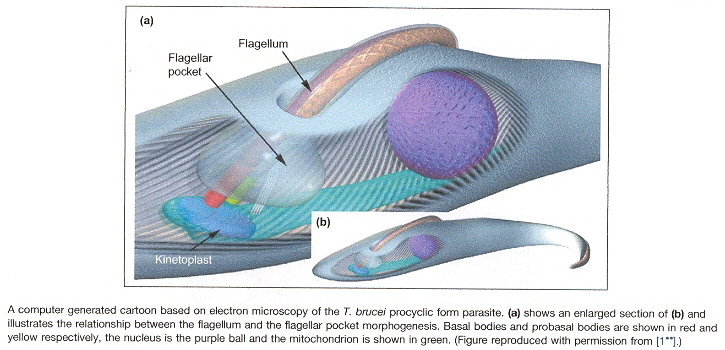
Flagella and cilia are organelles associated with kinetoplast mitochondria in protozoa as well as other eukaryotes. In order to understand the structure of the kinetoplast, abbreviations are common and may be found below. I addition, the basic structure is easily understood if examined in the figure below. Thus for example, 2K2N4BB means 2 kinetoplasts, 2 nuclei, 4 basal bodiesa, b.
Kinetoplast Abbreviationsa, b

T. brucei Kinetoplast-associated Organellesa
(Eukaryotic) Flagellum
Kinetoplast Structurec
Kinetoplast Diskd
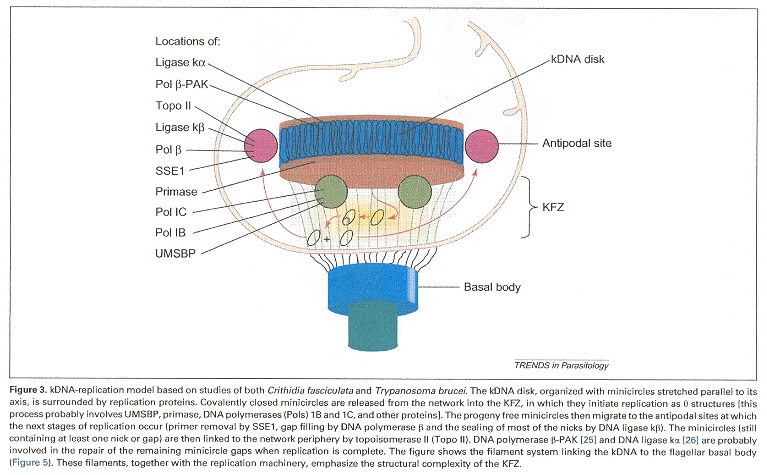
Morphology of the Kinetoplast Disk, Basal Body, antipodal structures, etc.e
More kDNA Kinetoplast Morphologye
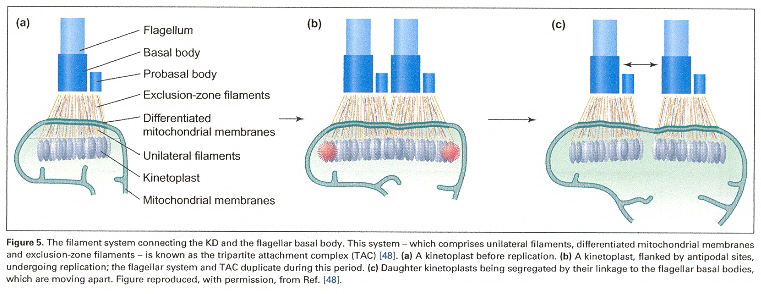
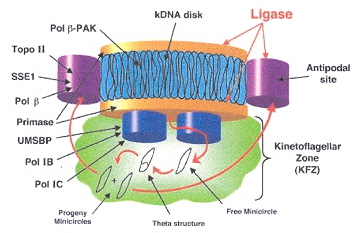
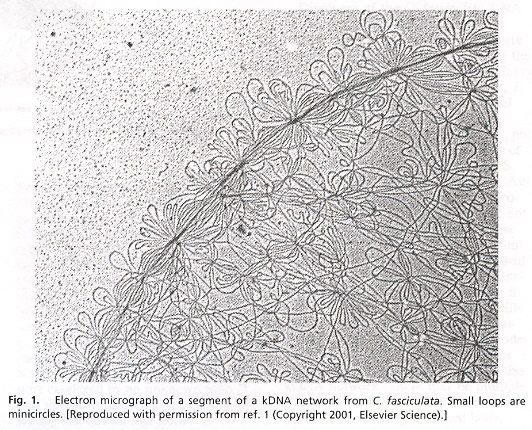
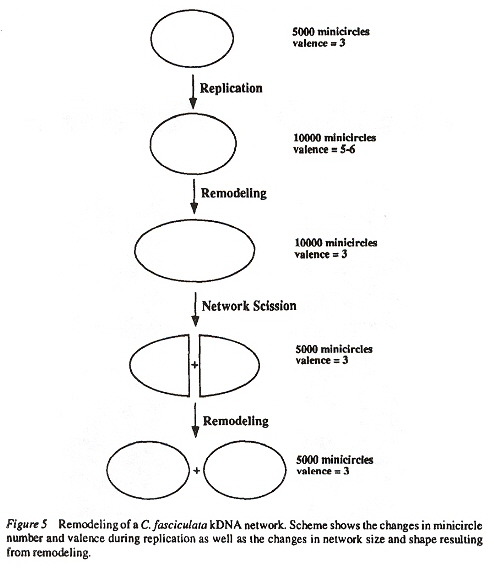
Examining the four figures above, one can see the Kinetoplast disk (as well as other structures, such as the basal body and the two antipodal sites which contain topoisomerases). Inside the kinetoplast disk may be found the network of thousands of DNA minicircles typically with a valence of 3 (the average is 3 minicircles linked together), as well as the few (25 to 50) maxicircles, all catenated together. This is the typical kinetoplast structure that can be found in Trypanosoma brucei, but older (from an evolutionary standpoint) is the Herpetomonas ingenoplastis which has a network in whick the circles are heterogeneous. The older Bodo caudatis has DNA circles which are not catenated. The more typical Trypanosoma brucei however, has a network that looks like the image below (minicircles have the appearance of chain mail, while maxicircles appear as rosettes). Note that in vivo, the minicircles are perpendicular to the kinetoplast disk with the minicircles parallel to the axis of the disk.
Catenated Network of Minicircles and Maxicirclesc
Minicircle Structured
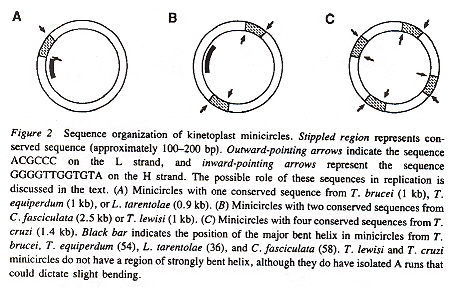
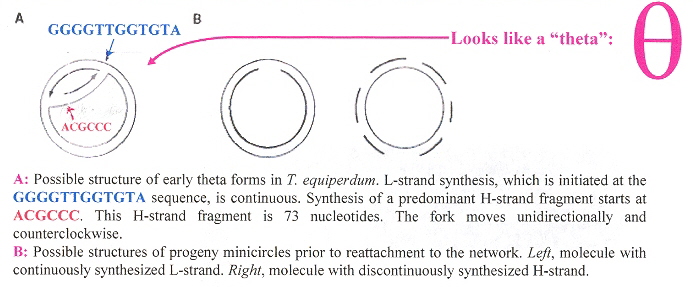
Closer biochemical analysis of the DNA minicircles reveals a very interesting structure (revealed in the figure above). The minicircles have an outer ring H-strand (the Heavy strand) and an inner ring L-strand (the Light strand). Each of these paired circles of DNA have a recognizable (conserved) area (some species have one, some two, some four such conserved structures). If there are two structures, they are 360°/2 = 180° apart, while if there are four such structures, they are 360°/4 = 90° apart. Each conserved area has conserved sequence blocks (CSBsg), typically named CSB-1, CSB-2, CSB-3. Each of these structures is identified by having the universal nucleotide base sequence of GGGTTGGTGTA on the H-strand (in CSB-3), and the universal sequence of nucleotide base sequence ACGCCC (in CSB-1) on the L-strand. See the figure below.
Universal Nucleotide Distiguished Sequencese
Antipodal Topoisomerase II Mediated Decatenation of the Network of DNA Circlesf
Kinetoplast Disk Antipodal Network Decatenationf
Kinetoplast Disk Spins to Randomly Select Separate Replicated DNA Minicircles to the Antipodesf
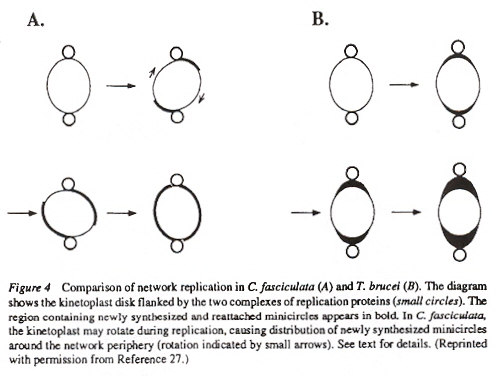
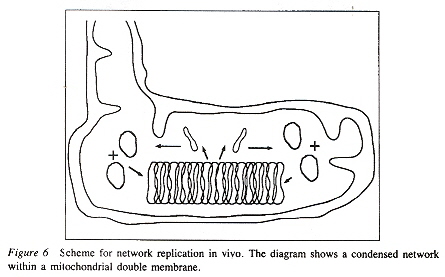
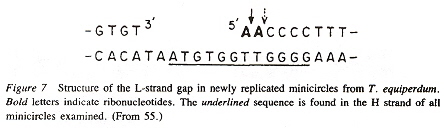
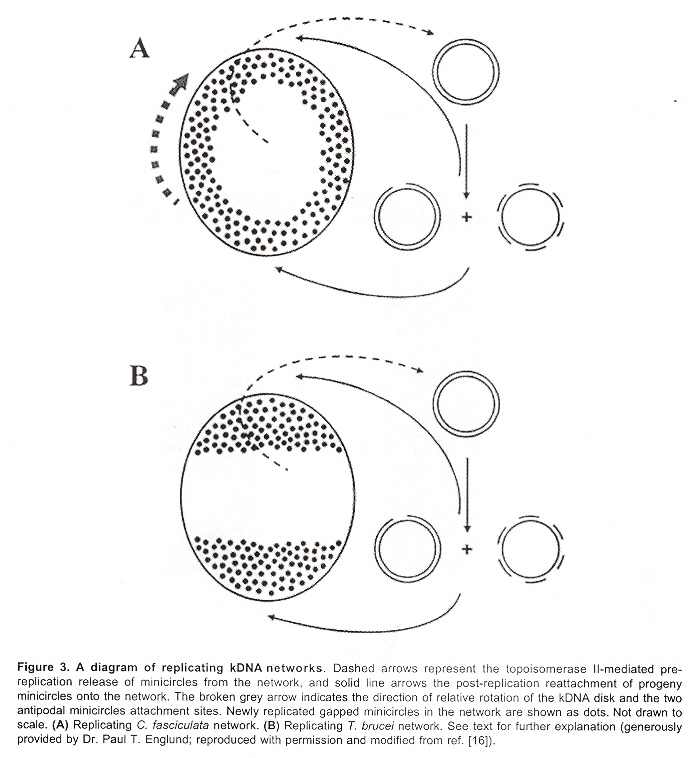
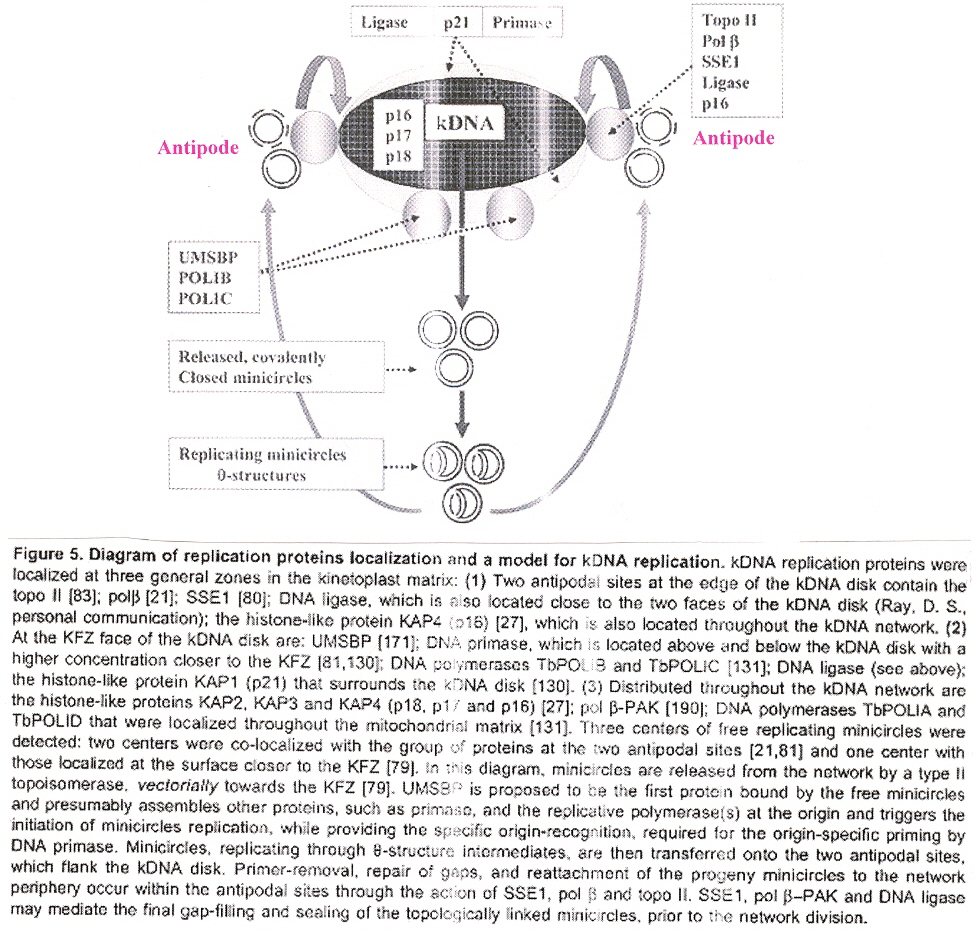
Maxicircles contain genes for rRNA, cytochrome b, and other proteins/catalysts, as well as ORFs (Open Reading Frames), but no tRNA in Kinetoplastids (Gillham reports tRNA in the H-strand in mtDNA of non-Kinetoplastids, and of course, exceptions in which tRNA has been found in Kinetoplastids exists, "Organelle Genes and Genomes", by N. W. Gillham, Oxford U.P., 1994, p. 54). A mitochondrial gene may be split into two fragments that are transcribed into twi nRNAs and translated into two polypeptides. It has been suggested that these two polypeptides are posttranslationally fused (for example, cytochrome oxidase is coded by the N-terminal part in mtDNA, while the C-terminal portion is encoded by nuclear DNAac. We now examine how these catenated kinetoplast DNA circles are replicated. Examine the three images above that deal with aspects of kDNA replication. The kinetoplast disk rotates (not in all protozoa kinetoplast replication) as the kDNA circles are replicated, randomly distributing copies so that one set of minicircles is at one antipode, its replicated minicircles being at the other antipode. To accomplish this, Topoisomerase II (type I cleaves a single strand of DNA, type II cleaves duplex strands) at each antipode decatenates the kDNA containing circles. During the replication process, the circles of kDNA become "nicked" and have "gaps", these "nicks" and "gaps" marking the circles that have been replicated. The complete repair of minicircle discontinuities is delayed until all minicircles have been duplicated. It appears that at this time, the basal body mediates the segregation of daughter progeny minicircles. Maxicircles replicate concurrently with the replication of minicircles. See the figure below. (Note that Saccharomyces has S type mitochondrial division, "Organelle Genes and Genomes", by N. W. Gillham, Oxford U.P., 1994, p. 111).
Network Replicationd
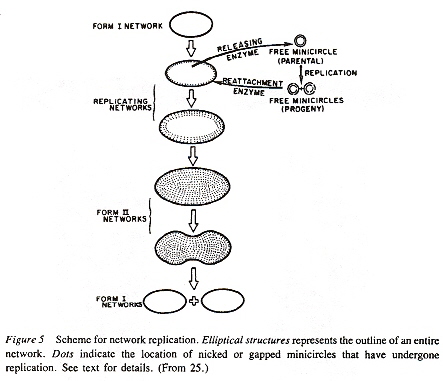
The kDNA replication process proceeds unidirectionally by theta ( θ ) structures.
kDNA Minicircle Replicationd
kDNA Maxicircle Replicationd
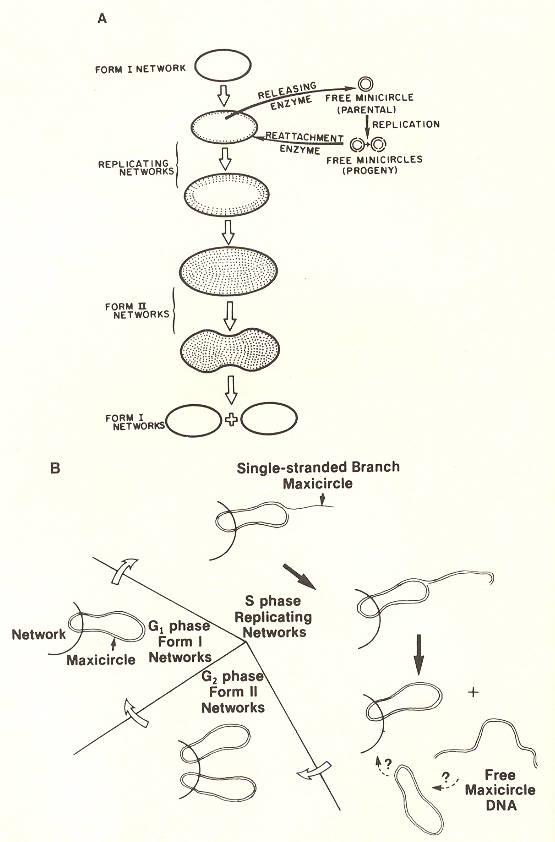
The theta structures form as the circles are replicated with the distingished universal minicircle H-strand and L-strand nucleotide bases being used to start replication of each outer and inner strand of DNA. As the Topoisomerase II acts to decatenate these circles, the circles are free, and will need a topoisomerase to re-catenate them and eventually, to reattach the catenated network to the paired (replicated) kinetoplasts. As the L-strand and H-strands are synthesized, the L-strand has a gap, of about ten nucleotides opposite the H-strand GGGGTTGGTGTA nucleotide sequence (named the universal minicircle sequence or UMS: this nucleotide sequence/gap is the origin of L-strand synthesis. When there are two or four conserved sequences (each with an identical gap, etc.), then synthesis starts at only one of the conserved sequences. Thus both the H-strand as well as the L-strand have distinguishing originating sequences where synthesis begins. As synthesis proceeds, the circle takes on a shape that looks like the Greek letter theta, hence the name θ replication. Maxicircle replication is also by theta replication, but is network-bound. Previously, it was thought that maxicircle replication was by rolling-circle replication, the replication fork proceeding unidirectionally. During minicircle replication, minicircles at the antipodes have a valence of 6 but after remodelling, the valence is reduced back to 3. The process of replication is complex, being composed of multiple steps (see the figure below). Note that analysis has shown (at least for T. brucei) that maxicircles are inherited from one parent, while minicircles are inherited from both parents.
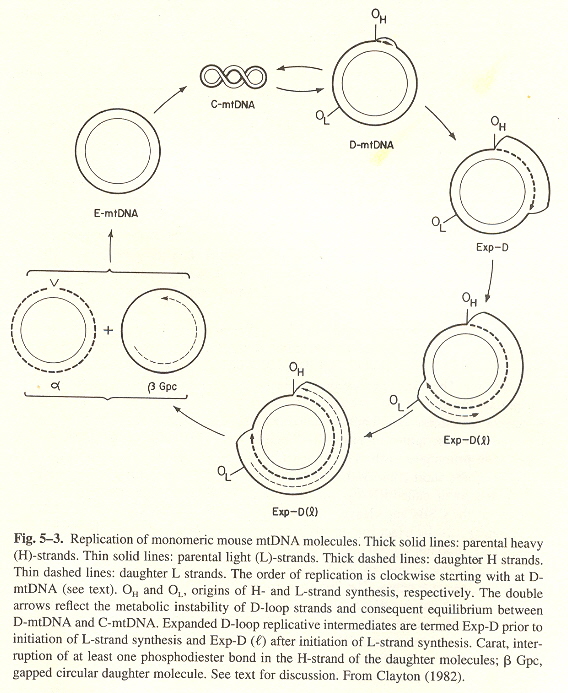
Animal mtDNA, as opposed to kDNA) replication is best described by mouse mtDNA, see the figure below. Replication on the H-srand starts at the OH origin, while replication on the L-strand starts later at the OL origin.
- H-strand triplex synthesis starts at OH
- H-strand replicates and continues about 2/3rds or the way, until OL is reached
- H-strand replication continues, while L-strand replication commences
- Two daughters are produced: α, and β Gpc (gapped circular), not yet completly synthesized
- The α daugthercompletes as a closed circle, the β Gpc daughter L-strand finishes synthesis
- Both daughters then become superhelical with about 100 negative turns: replication completed
"Organelle Genes and Genomes", by N. W. Gillham, Oxford U.P., 1994, pp. 117 - 118
Mouse mtDNA Replication
Remodelingf
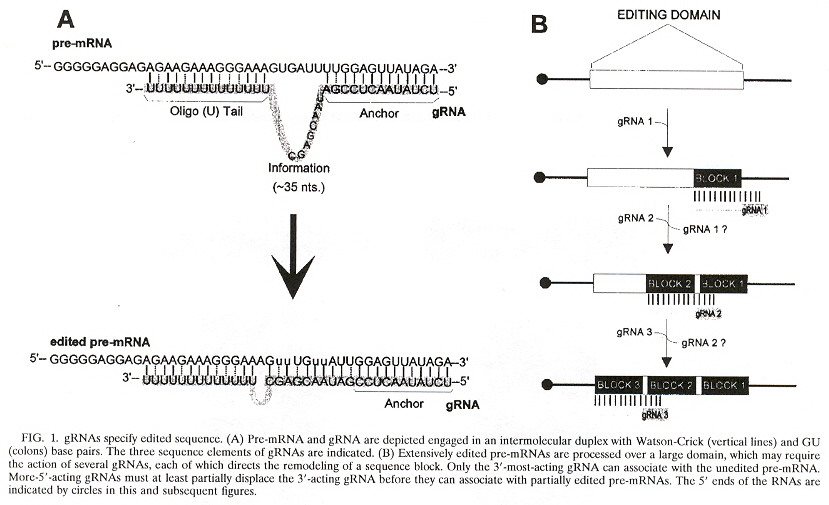
gRNA Editing to pre-mRNAh
gRNA Deletion and Insertion Editingh
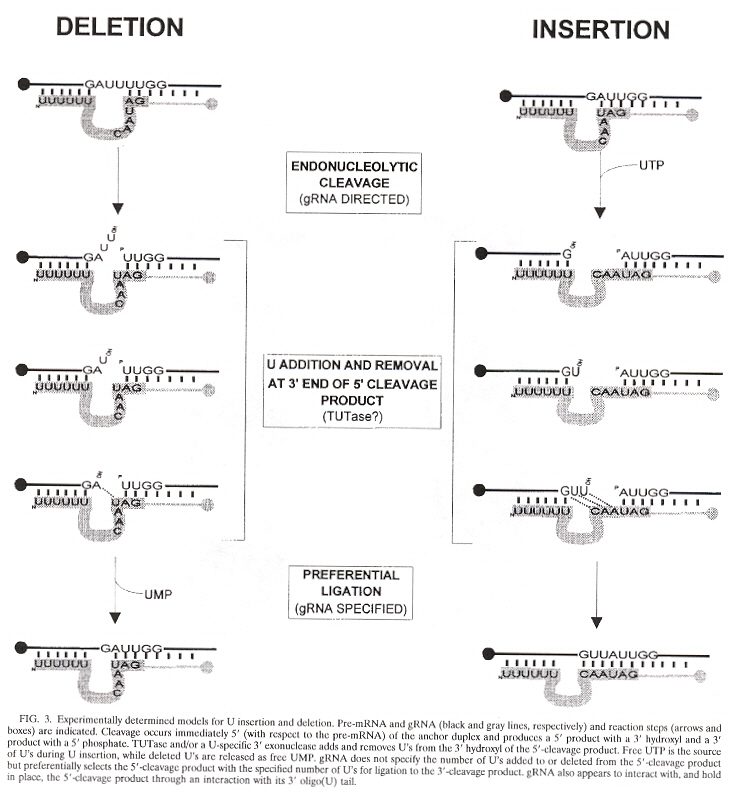
Details of gRNA Editing: U Insertioni
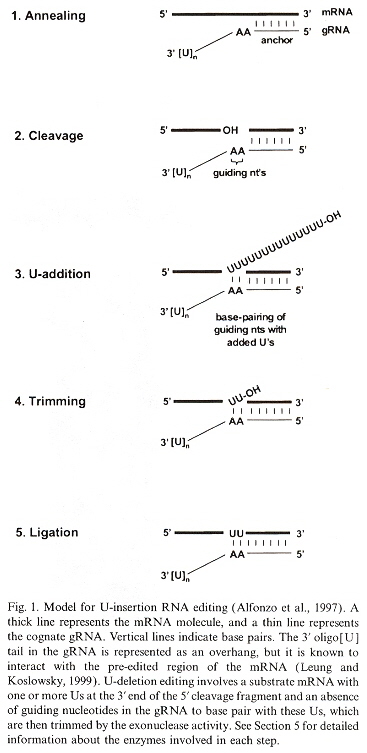
The maxicircles are "edited" to produce the final resultant nucleotide sequences. The editing consisting of the addition or deletion of uridines to the maxicircle corresponding pre-mRNA (creating the initiation and termination codons). The editing of the minicircles consists in modifying their corresponding pre-mRNA also, but uses the gRNA or guide RNA. This process of "editing" the pre-mRNA augments the usually understood central dogma. There typically is one or up to four gRNAs used for editing, but as would be expected, there are exceptions. T. brucei has more than 250 editing classes with at least 1,200 different gRNAs, while L. tarentolae has about 17 minicircle classes. The function of the minicircles is to encode gRNA (but maxicircles also encode some gRNAs). The edits fall into two classes:
- Base nucleotide insertion or deletion
- Base nucleotide modification
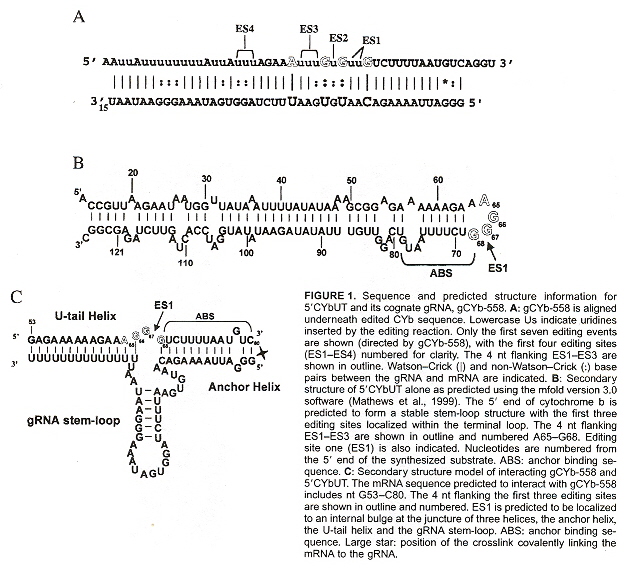
gRNAs specify the edited sequences in pre-mRNAs. gRNAs are typically 55 to 70 nucleotide bases in length. See the four figures above. Note that there is a 5' end gRNA anchor (that binds to the pre-mRNA anchor binding site, or ABS), followed by editing base sequence of about 25 to 35 bases that specifies where U insertion or deletion is to take place (called editing sites, or ES), terminated by the U tail of 5 to 24 bases at the gRNA 3' end. This U tail is associated with a gRNA endonuclease named terminal uridylyl transferase or tutase. More than half the pre-mRNA edits are U insertions with the final mRNA being twice as large as the original encoding gene.
Stretches of contiguous sequence which require one or more gRNAs are called editing domains. Most pre-mRNAs have a single editing domain, but T. brucei has multiple domains. Each domain is independently edited. pre-mRNAs are edited in the 3' to 5' direction. However, the exact timing of editing has not been determined. For example, it is possible that all deletion editing takes place, followed by all insertion editing (two peaks of editing activity have been observed). Another question: does gRNA remain associated with a region being edited until all sites are edited, or does gRNA dissociate after each site has been edited? What mechanism selects the order of edit sites? From the point of view of evolution, it appears that editing is more extensive in kinetoplastid groups that diverged early, than in those that evolved later.
ABS Anchor, ES Edit Sites, U-tailj
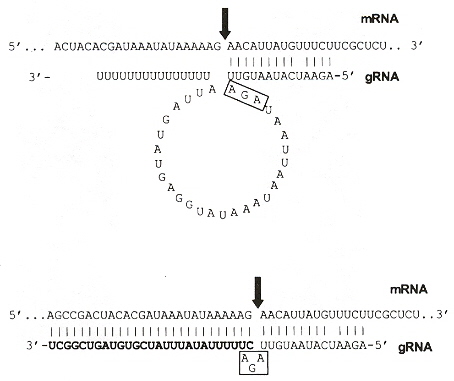
gRNA Editingi
a "Host-parasite interactions and trypanosome morphogenesis: a flagellar pocketful of goodies", by K. Gull, Current Opinion in Microbiology, Aug. 2003, 6, 4, 365 - 370
b "An evolutionarily Conserved Coiled-Coil Protein Implicated in Polycystic Kidney Disease Is Involved in Basal Body Duplication and Flagellar Biogenisus in Trypanosoma brucei", by G. W. Morgan, P. W. Denny, S. Vaughn, S. Goulding, T. R. Jeffries, D. F. Smith, K. Gull, M. C. Field, Molecular and Cellular Biology, May 2005, 25, 9, 3774 - 3783
c "Closing the gaps in kinetoplast DNA network replication", by M. M. Klingbeil, P. T. Englund, Proceedings of the National Academy of Sciences U.S., March 30 2004, 101, 13, 4333 - 4334
d "REPLICATION OF KINETOPLAST DNA IN TRYPANOSOMES", by K. A. Ryan, T. A. Shapiro, C. A. Rauch, P. T. Englund, Annual Reviews of Microbiology, 1988, 42, 339 - 358
e "Fellowship of the rings: the replication of kinetoplast DNA", by B. Liu, Y. Liu, S. A. Motyka, E. E. C. Agbo, P. T. Englund, Trends in Parasitology, Aug. 2005, 21, 8, 363 - 369
f "The Structure and Replication of Kinetoplast DNA", by J. Shlomai, Current Molecular Medicine, Sept. 2004, 4, 6, 623 - 647
g "THE STRUCTURE AND REPLICATION OF KINETOPLAST DNA", by T. A. Shapiro, P. T. Englund, Annual Reviews of Microbiology, 1995, 49, 117 - 143
h "RNA Editing in Kinetoplastid Protozoa", by K. Stuart, T. E. Allen, S. Heidmann, S. D. Seiwert, Microbiology amd Molecular Biology Reviews, March 1997, 61, 1, 105 - 120
i "Uridine insertion/deletion RNA editing in trypanosome mitochondria - a review", by A. M. Estévez, L. Simpson, Gene, 1999, 247 - 260
j "Interactions of mRNA amd gRNA involved in trypanosome mitochondrial RNA editing: Structure probing of an mRNA bound to its cognate gRNA", by S. S. Leung, D. J. Koslowsky, RNA Society, 2001, 7, 1803 - 1816
k "Effects of RNA Interference of Trypanosoma brucei Structure-specific Endonuclease-I on Kinetoplast DNA Replication", by Y. Liu, S. A. Motyka, P. T. Englund, The Journal of Biological Chemistry, Oct. 21 2005, 280, 42, 35513 - 355207
l "Biochemical and evolutionary aspects of anaerobically functioning mitochondria", by J. J. van Hellemond, A. van der Klei, S. W. H. van Weelden, A. G. M. Tielens, Philosophical Transactions of the Royal Society of London B, 2003, 358, 205 - 215
m "Kinetoplast DNA Minicircles Encode Guide RNAs for Editing of Cytochrome Oxidase Subunit III mRNA", by N. R. Sturm, L. Simpson, Cell, June 1 1990, 61, 5, 879 - 884
n "Universal Minicircle Sequence-binding Protein, a sequence-specific DNA-binding Protein that Recognizes the Two Replication Origins of the Kinetoplast DNA Minicircle", by K. Abu-Elneel, I. Kapeller, J. Shlomai, The Journal of Biological Chemistry, May 7 1999, 274, 19, 13419 - 13426
o "A single-stranded DNA binding protein binds the origin of replication of the duplex kinetoplast DNA", by D. Avrahami, Y. Tzfati, J. Shlomai, Proceedings of the National Academy of Sciences U.S.A., Nov. 7 1995, 92, 23, 10511 - 10515
p "Review Kinetoplast DNA", by P. Borst, J. H. J. Hoeijmakers, Plasmid, 1979, 2, 20 - 40
q "Kinetoplast DNA minicircles of phloem-restricted Phytomonas associated with wilt diseases of coconut and oil palms have a two-domain structure", by M. Dollet, N. R. Sturm, J-C. Ahomadegbe, D. A. Campbell, Federation of European Microbiological Societies Microbiology Letters, Nov. 27 2001, 205, 1, 65 - 69
r "Cruzrlla marina (Bodonina, Kinetoplastida): non-catenated structure of poly-kinetoplast DNA", by A. Zíková, M. Vancová, M. Jirků, J. Lukeš, Experimental Parisitology, July-Aug. 2003, 104, 3-4, 159 - 161
s "DNA TOPOISOMERASE PROTOCOLS, DNA Topology and Enzymes", Edited by M-A. Bjornsti, N. Osheroff, Methods in Molecular Biology, 1999, 94, 61 - 67 ("Isolation of Kinetoplast DNA", by T. A. Shapiro, V. A. Klein, P. T. Englund)
t "HETEROGENEITY IN SENSITIVITY TO CLEAVAGE BY THE RESTRICTION ENDONUCLEASES ECORI and HINDIII OF CIRCULAR KINETOPLAST DNA MOLECULES OF CRITHIDIA ACANTHOCEPHALI", by D. L. Fouts, D. R. Wolstenholme, H. W. Boyer, Journal of Cell Biology,, Nov. 1978, 79, 2 (Pt. 1), 329 - 341
u "Why kinetoplast DNA networks?", by P. Borst, Trends in Genetics, May 1991, 7, 5, 139 - 141
v "Kinetoplast DNA minicircles are inherited from both parents in genetic hybrids of Trypanosoma brucei", by W. Gibson, L. Garside, Aug. 1990, 42, 1, 45 - 54
w "MINI REVIEW KINETOPLAST DNA, MITOCHONDRIAL DNA WITH A DIFFERENCE", by K. Stuart, Molecular and Biochemical Parasitology, Oct. 1983, 9, 2, 93 - 104
x "KINETOPLAST DNA AND RNA OF TRYPANOSOME BRUCEI", by A. M. Simpson, L. Simpson, Molecular and Biochmical Parasitology, Dec. 1980, 2, 2, 93 - 108
y "Redox Potential Regulates Binding of Universal Minicircle Sequence Binding Protein at Kinetoplast DNA Replication Origin", by I. Onn, N. Milman-Shtepel, J. Shlomai, Eukaryotic Cell, April 2004, 3, 2, 277 - 287
z "Kinetoplast DNA of Bodo caudatus: a Noncatenated Structure", by S. L. Hajduk, A. M. Siqueira, K. Vickerman, Molecular and Cellular Biology, Dec. 1986, 6, 12, 4372 - 4378
aa "Mitochondrial and Nuclear Localization of Topoisomerase II in the Flagellate Bodo saltans (Kinetoplastida), a Species with Non-catenated Kinetoplast DNA", by I. Gažiová, J. Lukeš, The Jourrnal of Biological Chemistry, March 28 2003, 278, 13, 10900 - 10907
ab "Sequences within the 5' Untranslated Region Regulate the Levels of a Kinetoplast DNA Topoisomerase mRNA during the Cell Cycle", by S. G. Pasion, J. C. Hines, X. Ou, R. Mahmood, D. S. Ray, Molecular and Cellular Biology, Dec. 1996, 16, 12, 6724 - 6735
ac "Genomics and Evolution of Cellular Organelles", by M. S. Odintsova, N. P. Yurina, Russian Journal of Genetics, 2005, 41, 9, 957 - 967
It is proposed that kDNA ("Emergent Computation: Emphasizing Bioinformatics", p. 324) uses Topoisomerase II to form a complex, three-dimensional kDNA disk with catenated minicircles marked by nicks or gaps to show completion of replication to form the KFZ (kinetoflagellar zone) near the flagellar basal body. Of primary interest here is that a three-dimensional shape grammar could be used to model the processes described in this paper. The Appendix in "Emergent Computation: Emphasizing Bioinformatics" only discussed two-dimensional shape grammars.
Plasmids pVarkud and pMauriceville are circular plasmids that insert into mtDNA.
G-quartets in Kinetoplast DNA
"Sequences with high propensity to form G-quartet structures in kinetoplast DNA from Phytomonas serpens", by D. Sá-Carvalho, Y. M. Taaub-Cseko, Molecular and Biochemical Parasitology, June 1995, 72, 1-2, 103 - 109Kinetoplast maxicircles encode mitochondrial enzymes, and minicircles often encode gRNAs. As much as one third of these minicircle sequences is composed of G and T bases. These bases can form G-quartets with three different structural motifs:
- G4 (tetramolecular, parallel G-quartet quadruplexes)
- G'2 (bimolecular, antiparallel G-quartet quadruplexes)
- G4' (unimolecular, antiparallel G-quartet quadruplexes)
G-quartet Quadruplex Motifs
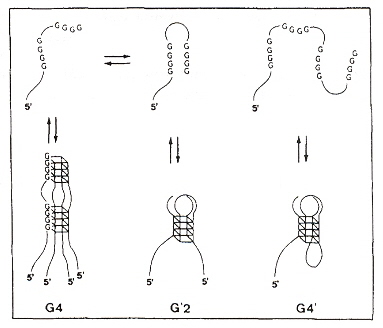
This paper describes G-quartet forming sequences found in kDNA minicircles that constitutes a polymerase arrest site. All three of the different structural motifs are found in vitro depending upon Na+ or K+ concentrations. These are listed in the table below. It is not argued that these quadruplex structures have been found in vivo, but that they are likely to exist would be a good experimental platform considering the high copy number as there are so many minicircles in kinetoplasts. Palindromic structures tend to form hairpins and this helps explain arrest-site properties.
G-quartet Quadruplex Oligonucleotide Polymerase-arrest site Sequences

A shape grammar for kDNA minicircles supporting multiple quadruplexes
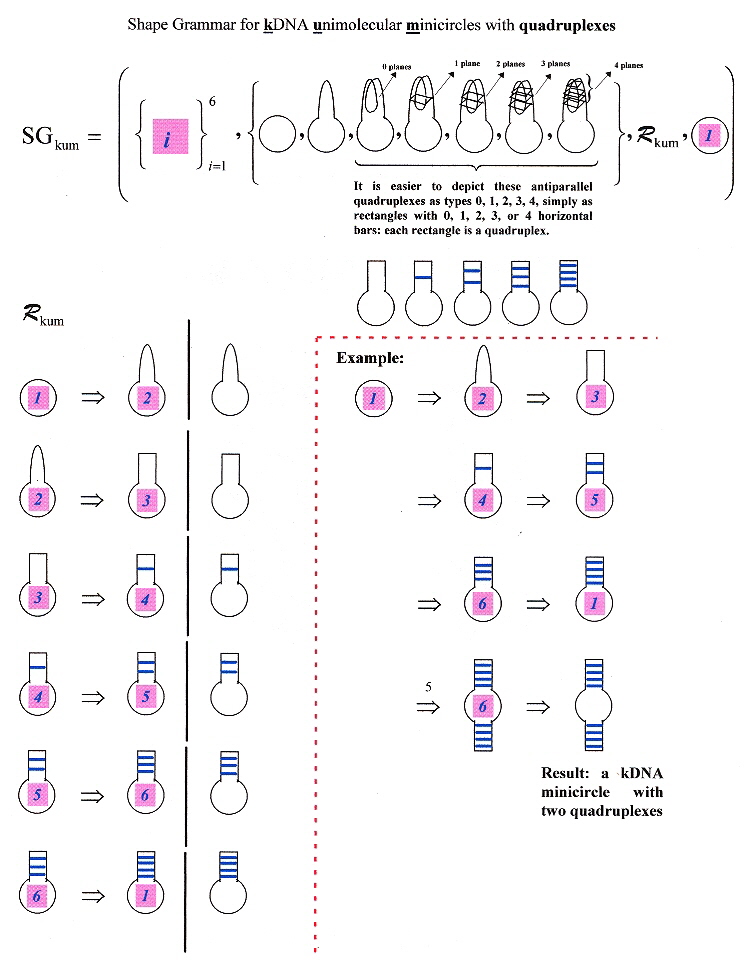
Background information about Plasmid Genomes in Kinetoplast DNA
Bacteria typically do not contain a nucleus, their DNA is located in cytoplasmic plasmids (typically circular in form, not linear, but there are exceptions as always: there are linear forms). Maize has a nucleus with DNA, as well as chloroplasts and circular-form plasmids (nuclear DNA can coexists with plasmid DNA). Plasmid DNA has been found in Neurospera intermedia mtDNA (thus plasmid DNA can integrate onto other DNA just as cpDNA and mtDNA can integrate with nuclear DNA and exist as nucpDNA and numtDNA). In fact, when the Kalilo plasmid DNA integrates with mtDNA, named kalDNA in Neurospora intermedia, this will result in the slow senescent death of Neurospera intermediaa.
It has been notedb that kalDNA originating from the kalilo plasmid (that normally resides in the cytosol), can insert itself into mtDNA in Neurospora intermedia. Thus was determined by using restriction endonucleases Eco RI and Hind III to open up the circular mtDNA into linear fragments. The insertion site in the mtDNA is located at an intron of a large rRNA gene. Initially it was not clear that the kalDNA was inserted into an ORF (Open Reading Frame) of the rRNA in the mtDNA. However, it was strongly felt that the kalDNA insert destroyed the intron ORF and affected the rRNA. The view was expressed that the kalilo insert increased the number of kalDNA modified mtDNA relative to the normal mtDNA, resulting in the progressive displacement of normal mtDNA. It was postulated that organisms with this defective mtDNA accumulated until a threshold was attained, whereupon the proportion of remaining functionally normal mtDNA was insufficient to support cell growth, thus leading to senescence. Upon closer examinationc, it was determined that kalilo plasmid DNA could integrate into several mtDNA sites, including at least three different positions within the mtDNA area that codes for the rDNA gene. Verification that kalDNA integrates into mtDNA was supported by crossing experiments that showed that kalDNA senescence follows a pattern of maternal inheritance (as expected, as mtDNA is inherited maternally).
The strains of Neirospora intermedia affected by kalilo plasmids are only found in Hawaiian islands. Another strain, Neurospora crassa with plasmid maranhar also shows senesence due to the progressive decline in cytochromes aa3 and b, but is found only in India. Maranhar also was demonstrated to be maternally inherited just as the mtDNA is. Does plasmid maranhar also integrate itself as marDNA into mtDNA? Great care was taken to ensure that marDNA in fact integrates into the mtDNA. The mtDNA were purified and treated with DNase I. DNase I will degrade any DNA outside the mtDNA as DNase I cannot access DNA enclosed within the organellsd. Results clearly showed that maranhar plasmid DNA in fact integrated as marDNA (in the mtDNA). Examination of electrophoretic mobility and restriction maps of maranhar distinguished maranhar from kalilo DNA. Both kalilo and maranhar were shown to have terminal inverted repeats. The same degenerative process that was observed to cause senesence with kalilo plasmids also operates with maranhar plasmids. The nucleotide sequence of the marinhar plasmid was examined, showing the inverted repeats, palindromic sequences, five ORFs, and potential start and termination codonse. Other examples of insertions of episomes have been found that integrate into mtDNA. Specifically, the S-1 and S-2 episomes in Zea maize f, g, h. Specifically, papers deal with Zea maize infertility. Maize cytoplasms that cause infertility may have the following classifications:
- N normal
- S USDA
- T Texas
- C Charrua
kalDNA

marDNA
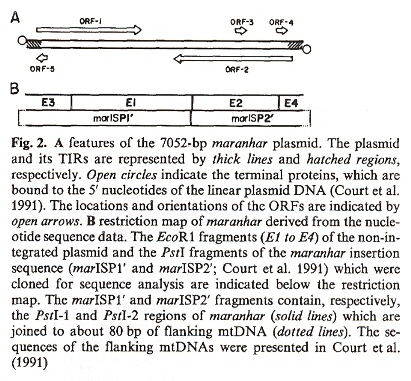
a "Organelle Genes and Genomes", by N. W. Gillham, Oxford U.P., 1994, pp. 247, 248)
b "Insertion of a Foreign Nucleotide Sequence into Mitochondrial DNA Causes Senescence in Neurospora intermedia", by H. Bertrand, B. S.-S. Chan, A. J. Griffiths, Cell, July 1985, 41, 3, 877 - 884
c "An Extrachromosomal Plasmid Is the Etiological Precursor of kalDNA Insertion Sequences, in the Mitochondrial Chromosome of Senescent Neurospora", by H. Bertrand, A. J. Griffiths, D. A. Court, C. K. Cheng, Cell, Dec. 5 1986, 47, 5, 829 - 837
d "A new senescence-inducing mitochondrial linear plasmid in field-isolated Neurospora crassa strains from India", by D. A. Court, A. J. F. Griffiths, S. R. Kraus, P. J. Russell, H. Bertrand, Current Genetics, Feb. 1991, 19, 2, 129 - 137
e "Genetic organization and structural features of maranhar, a senescence-inducing linear mitochondrial plasmid of Neurospora crassa", by D. A. Court, H. Bertrand, Current Genetics, Nov. 1992, 22, 5, 385 - 397
f "Unique DNA associated with mitochondria on the "S"-type cytoplasm of male-sterile maize", by D.R. Pring, C. S. Levings III, W. W. L. Hu, D. H. Timothy, Proceedings of the ational Academy of Sciences U.S.A., July 1977, 74, 7, 2904 - 2908
g "Mitochondrial DNA Rearrangements Associated with Fertile Revertants of S-type Male-Sterile Maize", by C. L. Schardl, D. R. Pring, D. M. Lonsdale, Cell, Nov. 1985, 43, 1, 361 - 368
h "The integrated forms of the S1 and S2 DNA elements of maize male sterile mitochondrial DNA are flanked by a large repeated sequence", by D. M. Lonsdale, R. D. Thompson, T. P. Hodge, Nucleic Acids Research, Aug. 11 1981, 9, 15, 3657 - 3668
Background information about Hydrogenosomes
"Evidence for Organelle Origin", by C. Q. Choi, The Scientist, March 28 2005, 19, 6, 25 - 26Hydrogenosomes generate molecular hydrogen, and are found in some ciliates, trichomonads, and chytridiomycete fungi. Finding DNA in hydrogenosomes has proven to be difficult. The more commonly held view is that hydrogenosomes either evolved from aerobic mitochondria that acquired an anerobic metabolism, or from endosymbionts capable of both aerobic and anerobic metabolism and that mitochondria descended from hydrogenosomes.
Using PCR with the anaerobic heterotrichous ciliate Nyctotherus ovalis a, b, DNA was found (below). It is not clear if the DNA found is of a circular or linear form.
- Mitochondrial complex I genes (nad2, nad4L, nad5, nad7)
- Mitochondrial complex I genes itegrated into the nuclear DNA
- Mitochondrial ribosomal protein genes (RPL2, RPL14)
- Mitochondrial complex II genes (Fp, Ip) itegrated into the nuclear DNA
- tRNA tyrosine gene
The hydrogenosomal bodies are bounded by a double membrane. Cristae-like projections (involved in electron transport) extend inwards from the inner membrane towards the center of the hydrogenosomal body. The hydrogenosome contains its own macronucleus and a micronucleus.
Nyctotherus ovalis Hydrogenosome
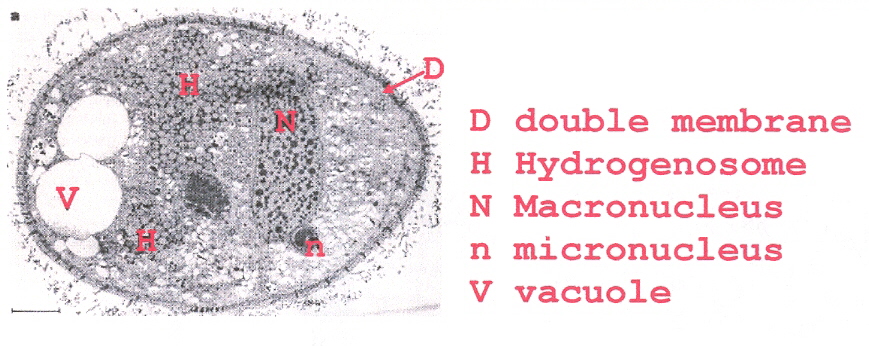
In addition, evidence was found that the hydrogenosomal organelle evolved from a mitochondrion, although this is not yet accepted by all researchers.
a "A hydrogenosome with a genome", by A. Akhmanova, F. Voncken, T. van Alen, A. van Hoek, B. Boxma, G. Vogels, M. Veenhuis, J. H. Hackstein, Nature, Dec. 10 1998, 396, 6711, 527 - 528
b "A hydrogenosome with pyruvate formate-lyase: anaerobic chytrid fungi use an alternative route for pyruvate catabolism", by A. Akhmanova, F. G. L. Voncken, K. M. Hosea, H. Harhangi, J. T. Keltjens, H. J. M. op den Camp, G. D. Vogels, J. H. Hackstein, Molecular Microbiology, June 1999, 32, 5, 1103 - 1114
Back to TopBackground information about Other Organelle DNA
There are other organelles besides the nucleus, chloroplasts, and kinetoplasts. Is DNA associated with other organelles? If DNA, RNA, proteins, etc. were to be found in other organelles, could such organelle DNA relocate between each other and perhaps even lend itself to linguistic/automaton studies?
-
"Evidence for the presence of DNA at basal body sites in Tetrahymena pyriformis", by Sir J. Randall F.R.S., C. Disbrey, Proceedings of the Royal Society of London, B, July 1965, 162, 989, 473 - 491
It has been noted that there are several reasons to suggest a search for DNA in the basal body of Tetrahymena pyriformis, including the fact that they multiply independently of the nucleus. The basal body is associated with flagella, cilia, and centrioles. Centrioles are of special significance in so far as they are associated with other fibrous structures such as the asters and spindles of mitosis, as well as the fibrils of the sperm tail.
I this study, it is concluded that the basal bodies of Tetrahymena pyriformis, strain S does contain DNA. Other studies seeking to find DNA associated with basal bodies do not find DNA. The authors of this paper did attempt to prevent contamination by DNA from sources other than basal bodies and to determine the precise times and conditions when DNA might best be detected. It was determined that the amount of basal body DNA is about 1% of the amount of DNA found in the cell nucleus. This amount of basal body DNA compares well with the 3.3% DNA found in Chlamydomonas reinhardii chloroplasts, and the 1% found in Neurospora crassa mitochondria.
-
"EVIDENCE FOR THE ASSOCIATION OF RNA WITH THE CILIARY BASAL BODIES OF TETRAHYMENA", by H. Hartman, J. P. Puma, T. Gurney Jr., Journal of Cell Science, 1974, 16, 241 - 259
A new procedure is used that provides better and more stable isolation of Tetrahymena pellicles (basal bodies). Other researchers found evidence of nucleic acid in basal bodies. The newer methods reported in this paper were able to detect fluorescence independently of the stage of the cell cycle (not true previously). Given the experimental evidence, the authors conclude that basal bodies contain a single-stranded nucleic acid: RNA generated from the cellular nucleus, where the RNA is held in rigid configuration by proteins.
-
"Basal Body/Centriolar DNA: Molecular Genetic Studies in Chlamydomonas", by J. L. Hall, Z. Ramanis, D. J. L. Luck, Cell, Oct. 6 1989, 59, 1, 121 - 132
In Chlamydomonas reinhardtii, the basal bodies at the root of both of its flagella are duplicated during mitosis, and are then repositioned at the spindle poles and then serve as centrioles, thus the connection between basal bodies and centroles (in some organisms). Analysis found circular DNA located in the basal body organelle. Evidence supports the view that each basal body contains circular DNA. The exact relationship between nuclear DNA, chloroplast DNA, mitochondrial DNA, and basal-body DNA has not yet been fully determined.
-
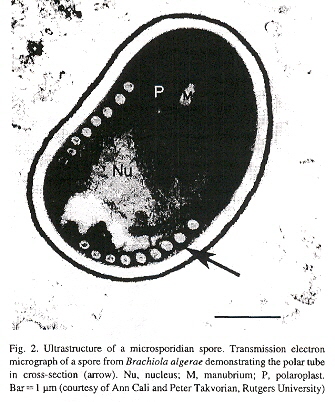
"The microsporidian polar tube: A highly specialized invasion organelle", by Y. Xu, L. M. Weiss, International Journal for Parasitology, 2005, 35, 941 - 953
Microsporidia possess a polar tube organelle. In the spore, the polar tube is connected to the anterior end, then coils around the sporoplasm. Under appropriate environmental stimulation the polar tube rapidly discharges out of the spore, piercing a cell, serving then as a conduit for sporoplasm passage into the new host.
Microsporidian Spore
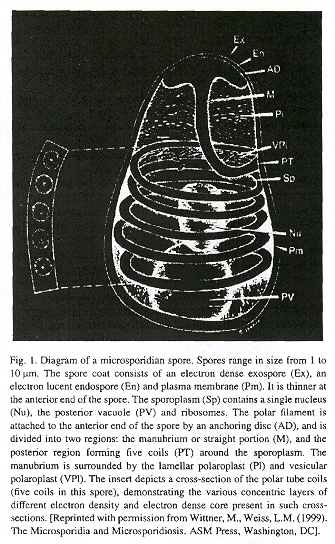
Microsporidian Spore
If all the information is gathered, what is found is that organelles remain poorly understood: How did they evolve, what is their relation to the nucleus, etc.? Centrioles, golgi bodies, basal bodies, flagella, cilia, etc. may not contain DNA, and certainly whatever is finally found may vary by organisms. However, hereditary templating (copying) information can be passed in other ways than DNA, such as RNA, proteins, etc. As such, no matter what the templating mechanisms, it is quite possible that some of these mechanisms may be amenable to linguistic-patterns of organization.
Background information about Plasmid DNA a
Dr. Esther Lederberg, Lecture at Kanazawa Medical School, Japan, 1962

Just as DNA from mitochondria (mtDNA) can integrate itself into nuclear DNA as numtDNA, it appears as plasmid DNA (extranuclear DNA) can integrate itself into nuclear DNA, but it is named an "episome" in this caseb.
When two plasmids cannot be stably maintained in the same bacterial cell, then they are said to be from different classes. Thus compatibility depends upon ability to replicate. There are a number of plasmid incompatibility classes. The method of determining mutual campatibility is to introduce plasmid A into plasmid strain B, usually measuring the results by antibiotic resistance. There can be three possible results:
- The only plasmids found in cell colonies are strain A, thus the strains are incompatible
- All resultant colonies contain both strains, thus the strains (plasmids) are compatible
- Both strains co-exist in some, but not all colonies: further testing is required
Twenty-four Incompatibility Classes
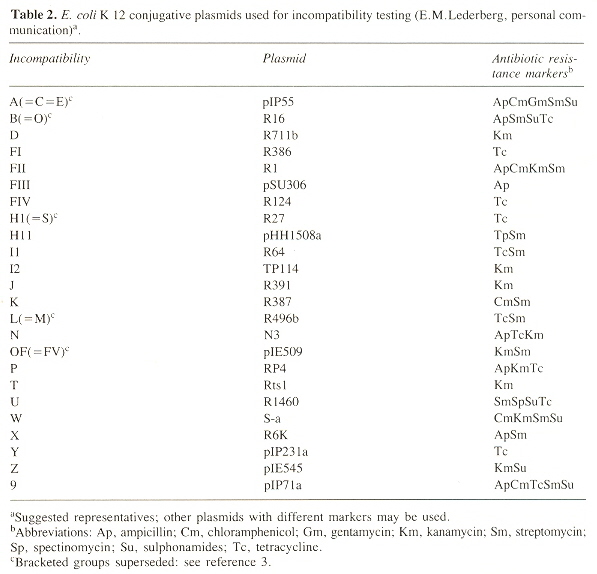
The names for the bracketed groups are as in the table: A superceds C and E; B superceds O; H1 superceds S; L superceds M; and OF superceds FV c
Thus at the outset, we are presented with a problem: almost anything said about plasmids might be true of one class, different in other classes. Thus the only approach to describe plasmids is to choose one of the most well known plasmids ("pT181") and use this as a paradigm for other plasmids. pt181 is a Staphylococcolal plasmid.
Drug resistance or R factors have outstanding functionsd:
- Hosts become resistant to antibacteriological agentse
- Host become genetic donors
The resistance to Shigella flexneri
- chloramphenicol
- tetracycline
- streptomycin
- sulfonamide
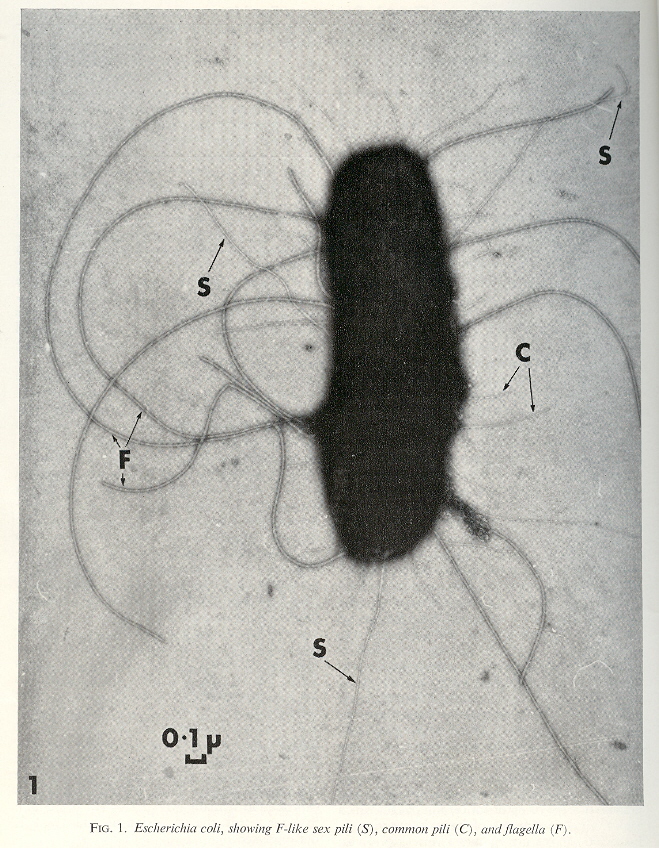
Resistance spread betweeen an enlarging number of different bacteria (by cell contact as well as phage), and to other antibiotics such as kanamycin, neomycin, penicillins, etc. Antibiotic resistance may be of less significance than the sex factor conferred by plasmid F or colicine factors (ColI, ColII). Due to the fertility factor F, not only does the cell become a donor, but it produces a new kind of filament not used for motility, the sex pilus. Note that conjugation reqires an F+ (with an F pilus) with an F– (without an F pilus).
F-originated Sex Pilus
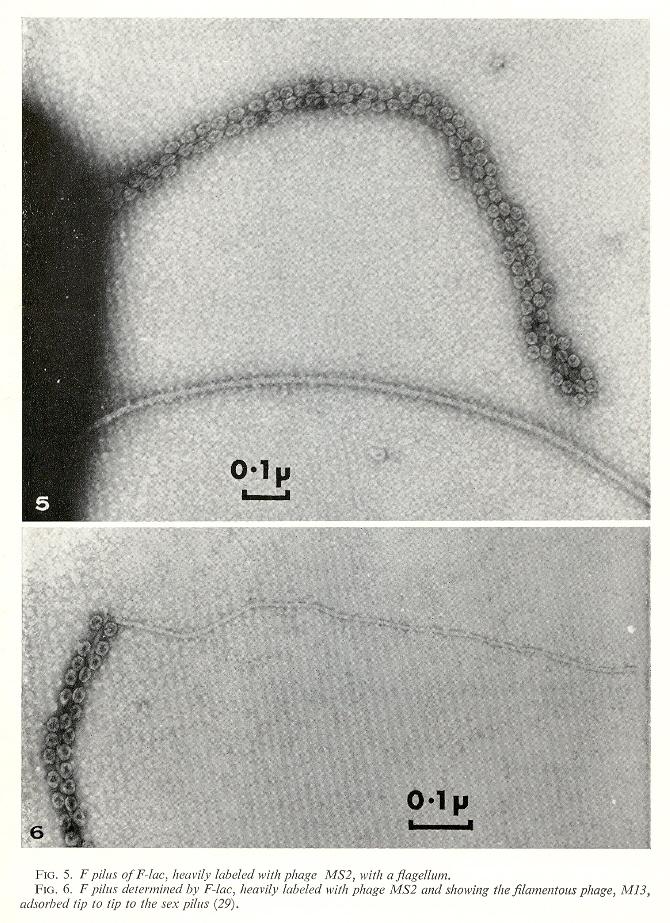
There are F-specific bacteriophages that target bacteria with F-pili. We can get an idea of their size by examining the next figure. There are different kinds of phages: isometric phages with RNA, and filamentous phages with single-stranded and double-stranded DNA.
Phages
There are in fact many different plasmids that may be classified in different ways. One way is to classify plasmids by their incompatibility reactions. Plasmids may be classified by their pili, and the effects of phages upon their pili. Specifically, can different plasmids coexist stably in the same bacterial strain? Three functions are especially significant:
- replication
- pilus formation regulation
- pilus structure
Classification by incompatibility reactions, including plasmid-specific phage, thermosensitive transfers, etc.
Plasmid Pili Classes
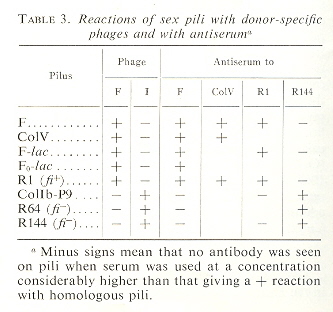
Plasmid Incompatibility Classes

Ira Herskowitz coined the term "cassette" that applies to a regulatory block of genetic information that can be silent when in storage at a silent locus but is expressed only when at a specific locus, see "CELLULAR DIFFERENTIATION, CELL LINEAGES, AND TRANSPOSABLE GENETIC CASSETTES IN YEAST", by I. Herskowitz, Current Topics in Developmental Biology, 1983, 18, 1 - 14. Private communication, E. Lederberg, 12/15/05
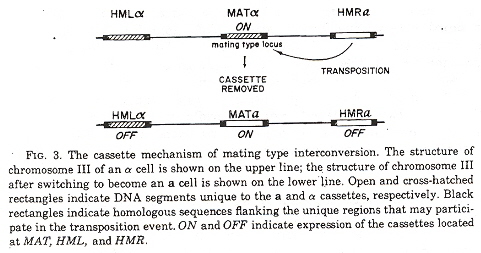
Genetic Casettes
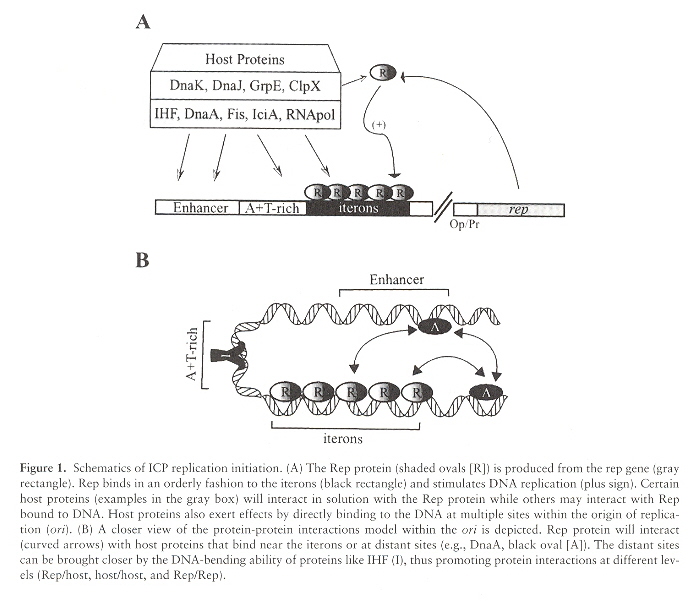
Plasmid DNA replication starts at the site named ori. Although in most cases there is only a single ori site, there can be more than one ori site. In the "typical" case, the ori site is recognized by Rep proteins that bind to DNA sequences called iterons. Plasmids are stable if their copy numbers are in equilibrium at a specidfic concentration. ori are composed of an Enhancer, an aditional region that is an A/T rich plasmid DNA area, followed by iterons, followed by a portion of the plasmid DNA named rep. The rep gene synthesizes Rep proteins. Rep proteins bind to the iterons. When Rep proteins bind to the iterons, plasmid DNA replication starts. This sequence may be modified by host proteins that can bind to the plasmid DNA which can facilitate DNA replication.
Ori (Plasmid Origens)
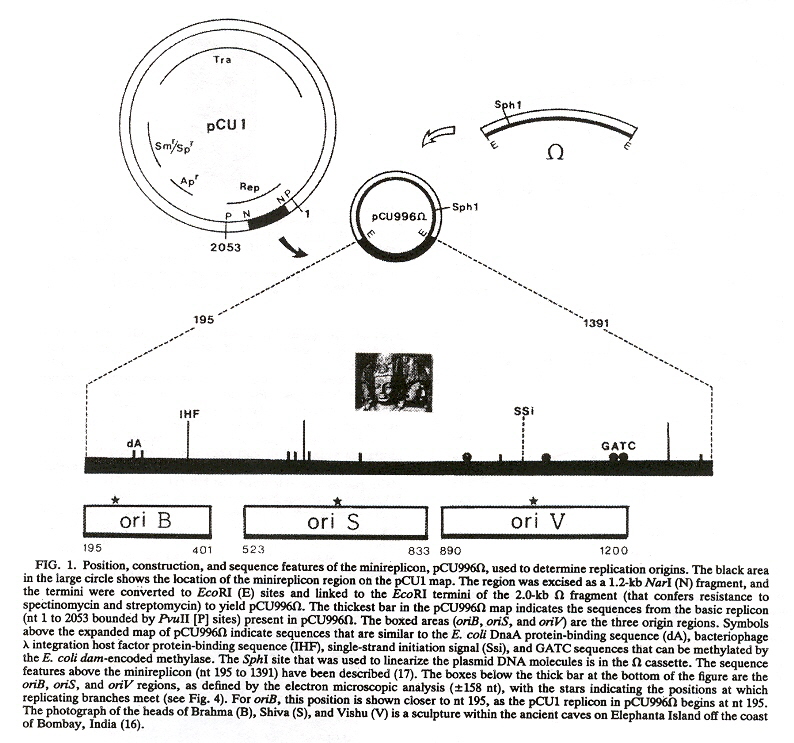
Plasmid Replication: ori
Most Gram-negative bacteria plasmids replicate by theta replication, while most Gram-positive bacteria plasmids replicate by rolling circle replication, and those that do tend to be small in sizef. Replication origins are dso (double-strand origin) or sso (single-strand origin). Plasmids that replicate by rolling-circle replication (RCR) may be grouped into at least seven families. Many RCR plasmids have the dso near the rep gene.
- pT181
- pC194/pUB110
- pE194/pLS1
- pSN2
- pGA1
- pG13
- pTX14-3
Seven major families of plasmids that replicate by Rolling-Circle
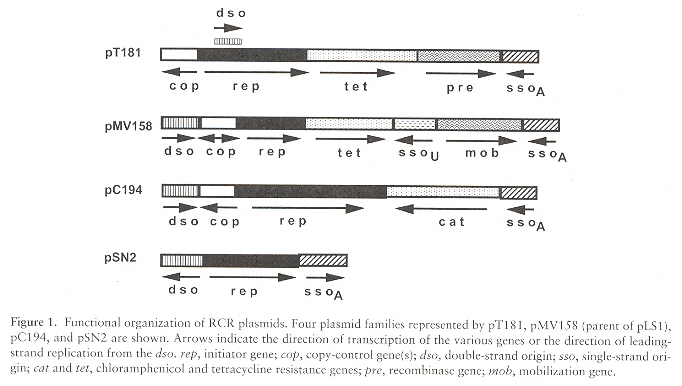
dso = (binding sequence region, nic sequence region). The binding sequence region is often contiguous or very close (14 to 95 nucleotide bases) to the nick site. Rep binds to the binding site. If the location of the binding sequence region and nic sequence region are artificially modified, the dso ceases to function. It is believed that the secondary and tertiary folding brings the binding sequence region into juxtaposition with the nic sequence region (even if these regions are not contiguous when viewed as a linear, primary sequence). A key feature of plasmids belonging to the same family is the conservation of the nic sequence that terminates replication. The bind sequence is typically 30 base pairs or less, of which about 20 base pairs are needed for sequence-specific Rep protein recognition. Of course, there are differences in these descriptions between pT181, pLS1, etc.
Using pT181 as our paradigm, the following are important pT181 plasmid classes g.
Staphylococcal Plasmids
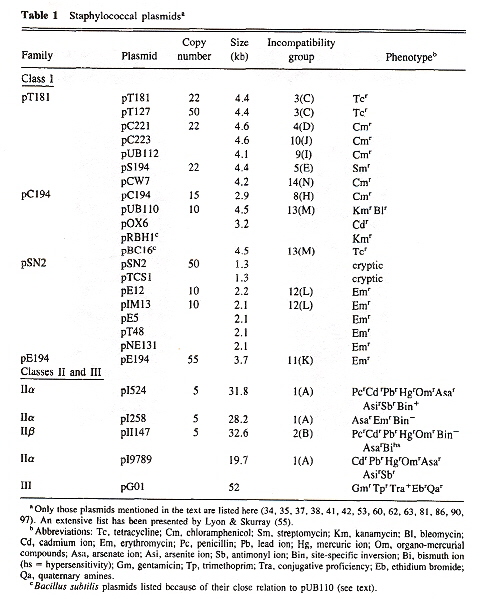
Each Ori (origin), and each Rep protein is commonly designated by a capital letter correspondig to the plasmid's incompatibility group (see the table above). Thus for classes I, II, and III, we have the following.
Staphylococcal plamid Ori and Rep proteins
The dso is in cruciform or hairpin-loop form. The Rep protein coded by the plasmid, interacts with the bind region of the dso. Then the dso is nicked by the Rep protein as the Rep protein covalently attaches to the 5' phosphate of the DNA (using tyrosine residue). PcrA helicase unwinds the nicked DNA, as ssDNA-binding protein SSB binds to the ss regions. DNA Pol III then synthesizes the leading-strand DNA (the replication fork progressing). Examine the figure below: an sso containing single-strand is synthesized at the far right. Note also at the far right, the Rep protein nicks this synthesized single-stranded plasmid DNA.The result (far right, bottom) is a replicated plasmid dsDNA, plus a single-stranded plasmid DNA with the sso located on a hairpin loop. The single-stranded plasmid DNA must now be replicated to synthesize the second copy of the plasmid dsDNA h.
Rolling Circle Replication, part I
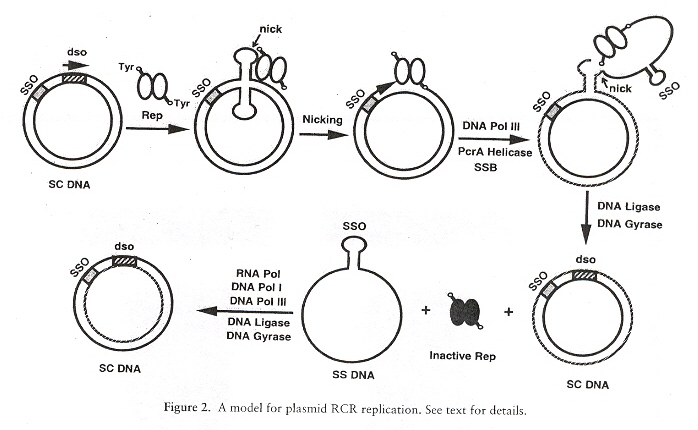
The figure above replicates the original double-stranded plasmid, plus the single-stranded plasmid DNA with the single-stranded origin sso. However, the single-stranded DNA containing the sso is then replicated to create a second double-stranded plasmid. Thus the overall process produces two plasmid dsDNA to complete the replication (see the figure below i).
Rolling Circle Replication, part II
While dso sequences are fairly well conserved within plasmid families, sso sequences are not necessarily conserved within the same family. Thus in one family, there are four different sso versions.
- ssoA
- ssoW
- ssoU
- ssoT
ssoA and ssoW function in their native host, while ssoU and ssoT function in many hosts. If sso's are deleted, reduced copy numbers and accumulation of ssDNA results. There are conserved sequences in sso found in hairpin loops. Similarly, there are conserved sequences in Rep proteins j.
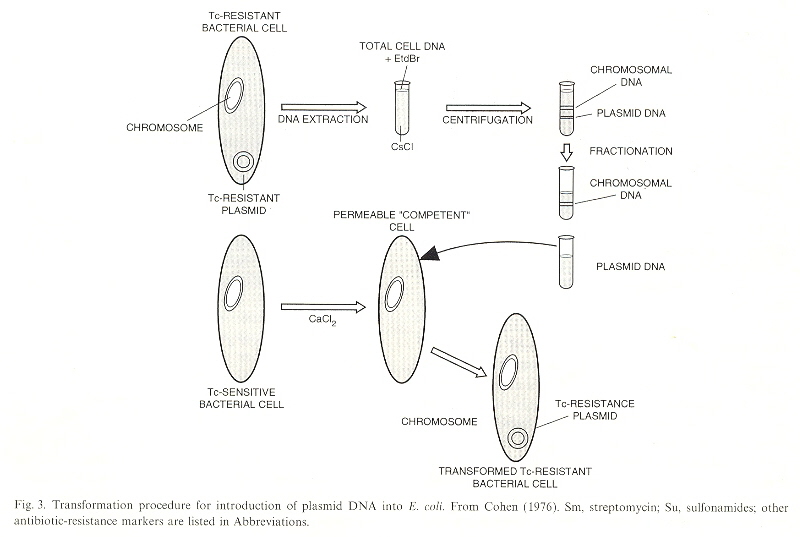
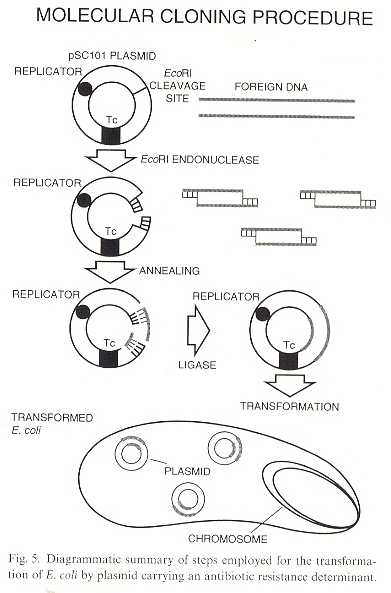
When DNA is extracted from Bacteria and centrifuged with CsCL, the result is that Chromosomal DNA and Plasmid DNA may be fractionated or separated. The plasmid DNA harvesed can be placed in the presence of other bacteria. In general, the bacterial membrane is not permeable to to DNA, but in the presence of CaCl2, the bacterial membrane is made permiable to plasmid DNA. Plasmid DNA, being promiscuous, could then modify the genome of the bacteria. As the plasmids could be used to introduce the same DNA to many bacteria at one time, this became known as cloning. Thus mutiple bacteria with the same genome could be used to test for different results introduced by antibiotic resistant plasmids, for example k. Plasmids are promiscuous, their DNA integrating to other DNA in mitochondria, chloroplasts, etc. in the same species, or different species. The plasmid DNA can be passed through transduction as well as conjugation.
Cloning: Duplicates
Cloning: Introducing Alien DNA
Interestingly, plasmids tend to locate at foci within the bacterial cell (usually equally spaced) to ensure distribution during cell division (however, this eqi-spacing takes place even when the cell is not dividing). Plasmids F, P1, and RK2 (compatible) position themselves within the cell. Positioning of the foci is at midcell and at the one-quarter and three-quarter positions. Each plasmid type appears to have its own unique subcellular address. After cellular duplication, the plasmids rapidly move to thier focal addresses l.
Plasmid Partitioning Foci
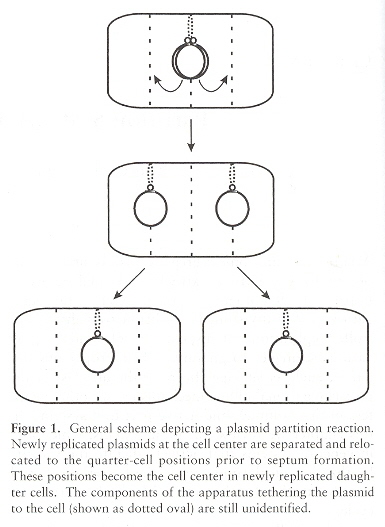
Lindenmeyer system for Plasmid Partitioning Foci
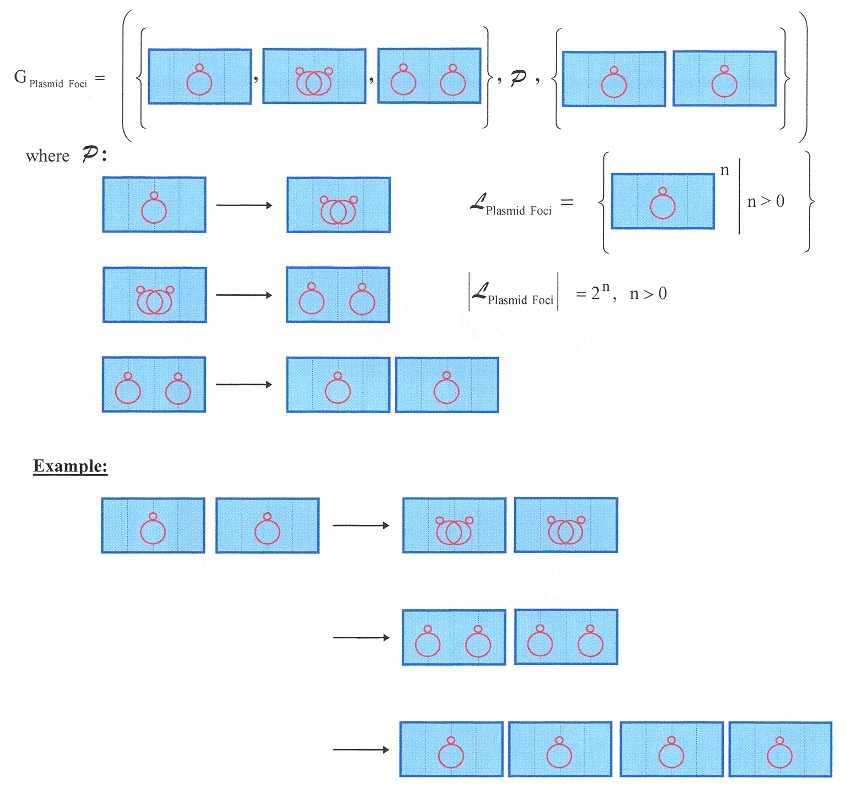
A small note concerning linear plasmids. Linear plasmid DNA have terminal telomeric structures. Actinomycetes and Borrelia differ in their telomeric structures, hence these structures may have evolved independently or through convergent evolution. The telomeric structures are as follows.
- Covalently closed hairpin loops (5' end joined to 3' end as with viroids), ex: Borrelia m
- Linear DNA with 5' end bound to a protein (protein-capped), ex: actinomycetes n
We shall not discuss replication of linear plasmids, this discussion being limited to a small note, and this shall end our discussion of plasmids, with the exception of one last note. A shape grammar can be constructed (not much more complicated than the "Lindenmeyer system for Plasmid Partitioning Foci" (above). A shape grammar can be constructed for promiscuous Maize chromosomes taking into account the synaptonemal complex, and chiasma formation during meiosis o.
a "Plasmid" coined by J. Lederberg, Private communication, E. Lederberg, 12/13/05
Also see "Cell genetics and Hereditary Symbiosis", by J. Lederberg, Physiological Reviews, Oct. 1952, 32, 4, 403 - 430
b Private communication E. Lederberg, 12/18/2005, in reference to "Molecular Structure of Bacterial Plasmids", by R. C. Clowes, Bacteriological reviews, Sept. 1972, 36, 3, 361 - 405.
"As presently used in bacteria, the term "extrachromosomal element" is synonymous with the term "plasmid,"..., p. 362
"Those plasmids which have been shown to be able to occupy a chromosomal site are referred to as episomes. p. 362. (chromosome is understood here to mean nuclear DNA, E. Lederberg).
"It may well be, therefore, that episomes are a special class of plasmids having an additional property of being able to integrate within certain host chromosomes,...", p. 363. (chromosome is understood here to mean nuclear DNA, E. Lederberg).
Compt rend. acad. sci., 1958, 247 - 154. F. Jacob and E. Wollman suggested that genetic elements which were optionally associated with the chromosomes of the cell be termed "episomes." Private communication, E. Lederberg, 12/14/05.
c Personal communication, E. Lederberg, 12/24/2005
d "Phylogenetic Relationships of Drug-Resistance Factors and Other Transmissible Bacterial Plasmids", by E. Meynell, G. G. Meynell, N. Datta, Bacteriological Reviews, Mar. 1968, 32, 1, 55 - 83
e Ibid. It is pointed out that that aside from R conferring antibiotic resistance, the ability to conjugate may also be donated. These factors are collectively referred to as RTF, or the "resistance transfer factor". The authors prefer the term the "sex factor", first applied by E. Lederberg to the F factor: plasmids capable of promoting nuclear chromosomal transfer. Transmissible drug resistance was discovered in Shigella flexneri as well as Salmonella typhimurium and other bacteria.
f "A novel rolling-circle-replicating plasmid from Pseudomonas putida P8: molecular characterization and use as vector", by R. Holtwick, A. von Wallbrunn, H. Keweloh, F. Meinhardt, Microbiology, 2001, 147, 337 - 344
g "STAPHYLOCOCCAL PLASMIDS AND THEIR REPLICATION", by R. P. Novick, Annual Reviews of Microbiology, 1989, 43, 538 - 563
h "Plasmid rolling-circle replication: recent developments", by S. A. Khan, Molecular Biology, 2000, 37, 3, 477 - 484
"Plasmid rolling-circle replication: highlights of two decades of research", by S. A. Khan, Plasmid, 2005, 53, 126 - 136
i "The Family of Highly Interrelated Single-Stranded Deoxyribonucleic Acid Plasmids", by A. Gruss, S. D. Ehrlich, Microbiological Reviews, June 1989, 53, 2, 231 - 241
j "Rolling circle-replicating plasmids from Gram-positive and Gram-negative bacteria: a wall falls", by G. del Solar, M. Moscosco, M. Espinosa, Molecular Biology, 1993, 8, 5, 789 - 796
"Replication and Control of Circular Bacterial Plasmids", by G. del Solar, R. Giraldo, M. J. Ruiz-Echevarría, M. Espinosa, R. Díaz-Orejas, Microbiology and Molecular Biology Reviews, June 1998, 62, 2, 434 - 464
"Plasmid rolling circle replication: identification of the RNA polymerase-directed primer RNA and requirement for DNA polymerase I for lagging strand sythesis", by M. G. Kramer, S. A. Khan, M. Espinosa, The European Molecular Biology Organization Journal, 1997, 16, 18, 5784 - 5795
k "Bacterial plasmids: their extraordinary contribution to molecular genetics (Recombinant DNA; extrachromosomal elements; transposons; gene regulation; DNA replication)", by S. N. Cohen, Gene, 1993, 135, 67 - 76 Table I in this paper (p. 71) was provided by E. Lederberg, private communication, 12/16/2005.
l "Compatible bacterial plasmids are targeted to independent cellular locations in Escherichia coli", by T. Q. Ho, Z. Zhong, S. Aung, J. Pogliano, The European Molecular Biology Organization Journal, April 2 2002, 21, 7, 1864 - 1872
m "Linear plasmids of Borrelia burgdorferi have a telomeric structure and sequence similar to those of a eukaryotic virus", by J. Hinnebusch, A. G. Barbour, Journal of Bacteriology, 1991, 173, 7233 - 7239
n "Interspecific transfer of Streptomyces giant linear plasmids in sterile amended soil microcosms", by J. Ravel, E. M. H. Wellington, R. T. Hill, Applied and Environmental Microbiology, 2000, 66, 529 - 534
o "Promiscuous Maize Chromosomes", by E. Martinez-Perez, G. Moore, Science, Jan. 2 2004, 303, 5654, 49 - 50
The plasmid copy number is significant. For example, it is possible that if a cell spilts, a daughter plasmid may not pass to a cell during replication, if the copy number is small. Thus P0 = 21-n, where "n" is the copy number, and "P" is the probability. Thus for example, plasmid pBR322 has n = 21, thus P0 = 21-21 = 2-20 = 1,048,576-1 (approximately 1 in a million).
Linear plasmids may have contributed DNA to plant mitochondrial genomes. Intact reading frames for RNA polymerase and DNA polymerase similar to those encoded by linear mitochondrial plasmids have been found in mtDNA.
Peptide Nucleic Acids (PNA) strongly bond to duplex DNA. A triplex is possible where an oligonucletide strand of PNA can bond to DNA, and then, mediated by a "linker", the PNA/linker can be bonded to a theraputic agent. Effectively, one can view this as an artificial, therapeutic PNA adduct. (review PNA Chiral-Chemistry). Currently, there are two major vectors to deliver therapeutics:
- viral
- non-viral
Any vector used to deliver therapeutics must balance safety vs. efficiency. Viral vectors are very efficient, but induce immune responses, and are limited to small therapeutic genes. It is hoped that PNA bound to therapeutics will provide high efficiency. The promise of gene-therapy is real: Dr. A. Fischer in Paris has cured several children with X-SCID (Severe Combined Immunodeficiency) using viral vectors, although some patients developed a leukaemia-like disorder.
PNA is bound to plasmid DNA . Linkers are attached to the PNA to provide conformal/steric space so as not to prevent the PNA from hybridising (binding) to the DNA. FEs (Functional Entities) are bound to the linker and deliver the therapeutic.
Plasmid/PNA Therapeutic Vector
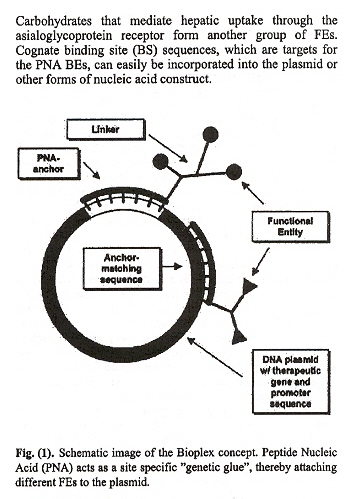
Viruses and Viroids
"Review: Identification of Structural Domains Involved in Astrovirus Capsid Biology", by N. K. Krishna, Viral Immunology, Spring 2005, 18, 1, 17 - 26
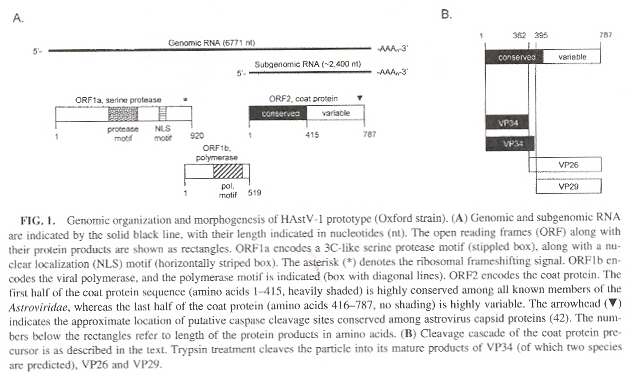
Astroviruses have two families (Mamastrovirus infect mammals, and Avastrovirus infect
birds). Astroviridae capsids have been filly sequenced in the following cases:
Astrovirus Genomic Organization
Astrovirus coat protein contains an assembly domain and a receptor domain as in the following figure.
Astrovirus Protein Coat Domains
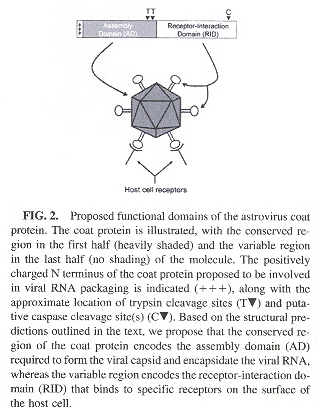
There are different kinds of viral genomes. The following is a partial description. However, only a very brief discussion about viruses and viroids will followa.
- Single-stranded DNA
- DNA: Linear
- DNA: Circular
- Double-stranded DNA
- DNA: Linear
- DNA: Circular
- RNA: circular
Circular viral DNA can be converted into linear form using restriction endonucleases (if not already linear). Hence linguistic methods that apply to either linear or circular DNA or RNA can be applied to the viral genome.
Viroidsb are a new class of pathogens (found in plants, but likely to be found much more widely in nature) which are distiguished from viruses by four very significant factors:
- The genome is of RNA (a single-strand of RNA)
- The absence of an encapsulating RNA protein coat
- Extraordinarily small size
- Viroids (PSTVd) do not code for any associated proteins
Size of Viroids, Viruses, and Bacteria
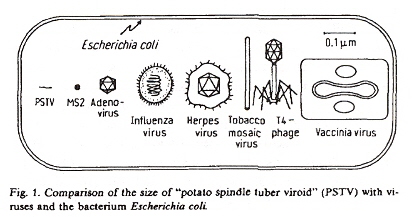
In the case of PSTVd (Potato Spindle Tuber Viroid), the RNA is a covalently closed ring (is circular, although non-infective linear-stranded RNA is present, too).
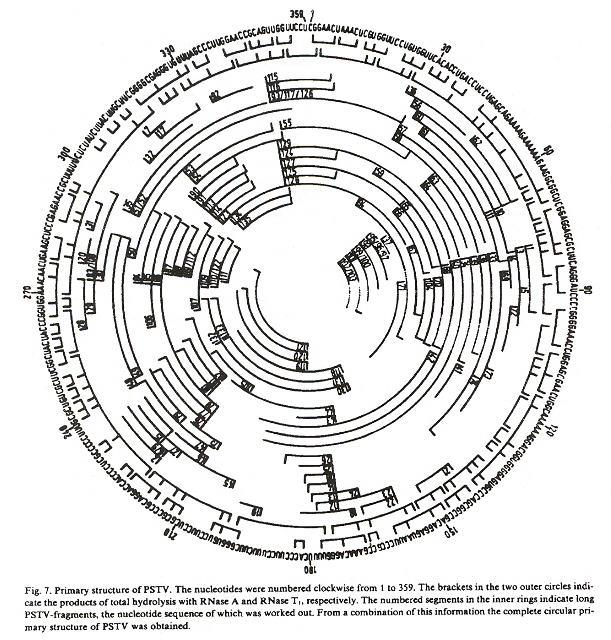
Overlapping fragments of the PSTVd genome have been put together to result in the following circular RNA.
Circular PSTVd RNA
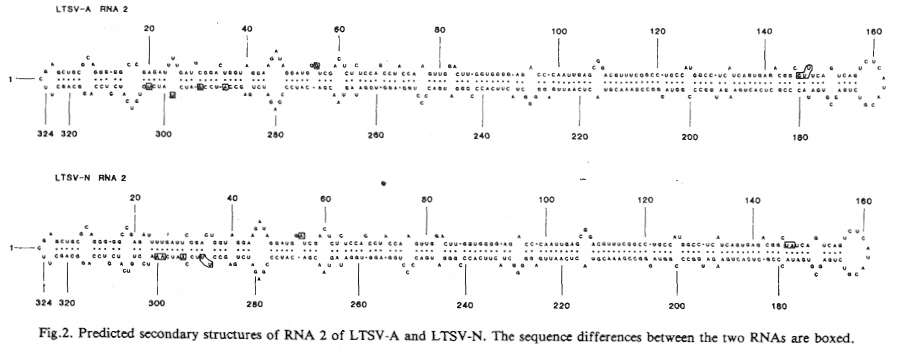
The authorsb point out that they redefine the usual meaning of tertiary structure. Tertiary structure usually means the three dimensional configuration, but is now redefined as the structure which is not the primary or secondary structure.
The secondary structure of viroids looks like the following. This is not for PSTVd, rather it is for LSTV-2 c, or Lucerne Transient Streak Virus. LTSV has two encapsulated single-strands of RNA, the larger RNA-1 is linear, but the smaller RNA-2 is a circular virusoid, or is viroid-like, and has a structure very similar to PSTVd. RNA-1 is capable of replication without RNA-2, thus the viroid-like RNA-2 helps in replication or is called a satellite viroid.
The RNA secondary structure tends to be extremely stable ex planta, and highly infectious. The potato spindle tuber viroid is rapidly spread through transencapsulation (encapsidated within potato leaf roll virus [PLRV]) f.
Typical Viroid RNA Secondary Structure
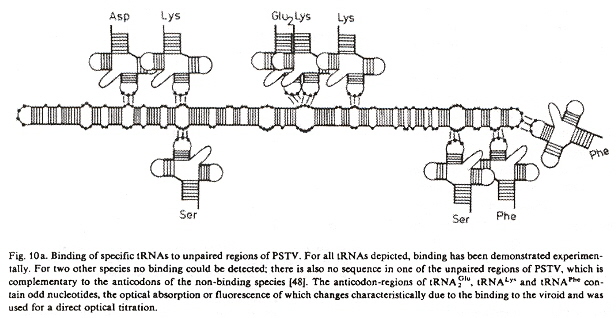
While viroids do not code for proteins, tRNAs do bind to loop unpaired base triplets (codons).
tRNA binding at viroid codons
Virusoids are circular viroid-like satellite RNAs that differ from viroids in that they are encapsidated. Thus some researchers consider viroids to be escaped satellite RNAs.
a "SpV1 AND SpV4, SPIROPLASMA VIRUSES WITH CIRCULAR, SINGLE-STRANDED DNA GENOMES, AND THEIR CONTRIBUTION TO THE MOLECULAR BIOLOGY OF SPIROPLASMAS", by J. Renaudin, J. M. Bové, Advances in Virus Research, 44, 429 - 463
b "Viroids: A Class of Subviral Pathogens", by H. J. Gross, D. Riesner, Angewandte Chemie International Edition in English, April 1980, 19, 4, 231 - 243
c "Comparative sequence and structure of circular RNAs from two isolates of lucerne transient streak virus", by P. Keese, G. Bruening, R. H. Symons, Federation of European Biochemical Societies Letters, Aug. 8 1983, 159, 1-2, 185 - 190
d "The strands of both polarities of a small circular RNA from carnation self-cleave in vitro through alternative double- and single-hammerhead structures", by C. Hernández, J. A. Daròs, S. F. Elena, A. Moya, R. Flores, Nucleic Acids Research, Dec. 11 1992, 20, 23, 6323 - 6329
e "Polydanviridae - A Proposed Family of Insect Viruses with Segmented, Double-Stranded, Circular DNA Genomes", by D. B. Stoltz, P. Krell, M. D. Summers, S. B. Vinson, Intervirology, 1984, 21, 1 - 4
f "Investigating the specificity of real-time PCR assays using synthetic oligonucleotides", by N. Boonham, T. Fisher, R. A. Mumford, Journal of Virological Methods, Dec. 5 2005, 130, 1-2, 30 - 35
"Viroids", Editors: A. Hadidi, R. Flores, J. W. Randles, J. S. Semancik, CSIRO Publishing, Australia, 2003
At this point in time, viroids have only been found in plants a. Viroids are covalently bound circular RNA, that differ from viruses and viral satellite RNA in that they exist in unencapsulated form and do not code for any proteins (they utilize host proteins for most biological functions such as replication, thus may be viewed as minimal parasites). While viroids tend to have ill-effects such as stunted growth, nevertheless there can be positive consequences too (as in stunted growth resulting in smaller plants, more easily sprayed with insecticides and fungicides, requiring less irrigation, easier fruit-gathering, premature-ripened fruit, increased yield, enhanced vigor and longevity of trees, etc). While viroids may lack symptoms of infection, they may thus serve as potential reservoirs of disease. Replication in the case of POSPIVIROIDAE takes place in the nucleus, while in the case of AVSUNVIROIDAE replication takes place in chloroplasts (no other organelles have been observed to be involved in replication, it is not known if replication can take place in other plastids). Replication in the POSPIVIROIDAE family is assumed to follow a single asymmetric rolling–circle; step that eventuates in polarity conservation. For the AVSUNVIROIDAE family, it is by symmetric rolling–circle; (that requires two replicative steps to preserve polarity). A review of polarity is in the paper "Identification of a Peach Latent Mosaic Viroid Hairpin Able To Act as a Dicer-Like Substrate", by Landry (below). Transmission is often by mechanical abrasion, grafts, but arthropods such as aphids have been efficient transmitters as well. Low temperatures and lack of light tend to arrest symptoms, while high temperatures and light tend to facilitate symptoms. One viroid tends to provide temporary cross-protection and interference from additional viroids, likened to immunization. There is implication of RNA–directed DNA methylation (RdDM) by viroid cDNA into the host genome, thus RNA gene-silencing as a novel form of epigenetic regulation, but explicit experimental evidence is still lacking.
Symmetric & Asymmetric Rolling-circle Replication
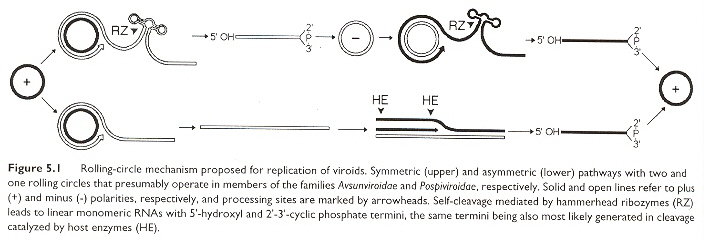
Viroid Negative-Polarity Rolling-circle Replication
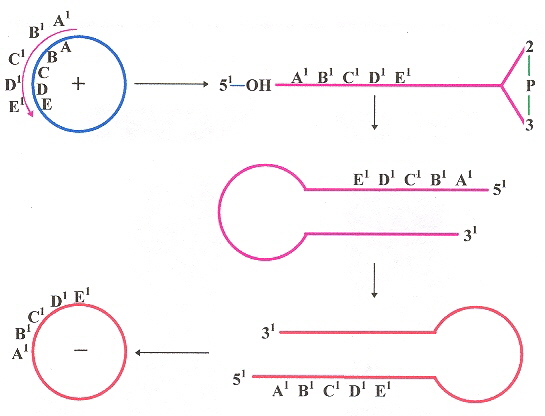
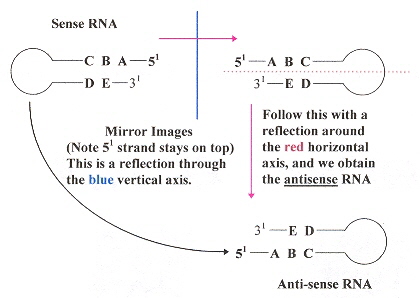
Note: The "negative" sense anti-RNA are base-complements, thus cRNA of the "plus' sense RNA
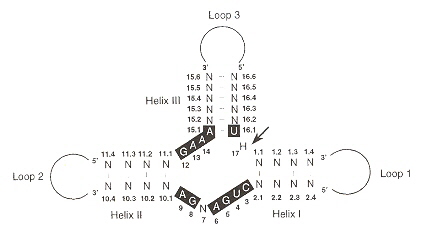
Viroids range in size from 246 bases to 399 bases. The POSPIVIROIDAE family has a rod-like secondary structure without bifurcations, composed of five domains. The rod-like structures in the TL domain may often undergo transitions into branched structures. The "UCCR" region can form hairpins (see below) but ae non-self-cleaving. AVSUNVIROIDAE on the other hand, are self-cleaving (by hammerhead structures).
- TL (Terminal Left, or T1)
- P (Pathogenic)
- C (Central, with upper central conserved region, UCCR; and lower central conserved region, LCCR)
- V (Variable)
- TR (Terminal Right, or T2)
As noted previously, the Avsunviroidae family of viroids have base sequences that can take on a hammerhead conformation, capable of self-cleavage ribozyme activity useful diring replication in either plus or minus polarities. Such a ribozyme conformation is composed of three helices and a catalytic core of 11 bases.
Viroid Self-Cleaving Ribozyme Conformation
Aside from viroids, there also exists viroid-like satellite RNAs (VL-satRNAs) are very similar to viroids, but depend upon a "helper" virus. satRNAs re distinguished by a v (circular) or s (linear) prefix to the helper virus. Thus a few examples follow.
| Helper Virus | VL-sat RNA | Size | Genus of | Encapsidated |
|---|---|---|---|---|
| Abbreviation | (nt) | Helper Virus | RNA Form | |
| Lucern transient streak virus | vLTSV | 324 | Sobemovirus | Circular |
| Subterranean clover mottle virus | vSCMoV | 332 | Sobemovirus | circular |
| 388 | ||||
| Velvet tobacco mottle virus | vVTMoV | 366 | Sobemovirus | circular |
| Solanum nodiflorum mottle virus | vSNMV | 377 | Sobemovirus | circular |
| Rice yellow mottle virus | vRYMV | 220 | Sobemovirus | circular |
| Tobacco ringspot virus | sTRSV | 359 | Nepovirus | linear |
| Chicory yellow mottle virus | sCYMV | 457 | Nepovirus | linear |
| Arabis mosaic virus | sArMV | 300 | Nepovirus | linear |
| Cereal yellow dwarf virus–RPV | sCYDV–RPV | 322 | Polerovirus | linear |
Using the International Committee on Taxonomy of Viruses (ICTV) guidelines, known viroids are as follows (using three criteria).
- Presence or absence of CCR
- Presence or absence of hammerhead ribozymes
- Replication by asymmetric or symmetric rolling-circle
| FAMILY | GENUS | SPECIES | Number of Variants |
|---|---|---|---|
| POSPIVIROIDAE | POSPIVIROID | PSTVd (potato spindle tuber)45 | |
| TCDVd (tomato chlorotic dwarf) | 1 | ||
| MPVd (mexican pepita) | 9 | ||
| TPMVd (tomato planta macho) | 1 | ||
| CSVd (chrysanthemum stunt) | 10 | ||
| CEVd (citrus exocortis) | 33 | ||
| TASVd (tomato apical stunt) | 3 | ||
| IrVd (iresine I) | 1 | ||
| CLVd (columnea latent) | 3 | ||
| HOSTUVIROID | HSVd (hop stunt) | 82 | |
| COCADVIROID | CCCVd (coconut cadang–cadang) | 7 | |
| CTiVd (coconut tinangaja) | 2 | ||
| HLVd (hop latent) | 10 | ||
| CVd–IV (citrus IV) | 1 | ||
| APSCAVIROID | ASSVd (apple scar skin) | 4 | |
| CVd–III (citrus III) | 22 | ||
| ADFVd (apple dimple fruit) | 1 | ||
| GYSVd 1 (grapevine yellow speckle 1) | 35 | ||
| GYSVd 2 (grapevine yellow speckle 2) | 1 | ||
| CBLVd (citrus bent leaf) | 7 | ||
| PBCVd (pear blister canker) | 11 | ||
| AGVd (Australian grapevine) | 1 | ||
| COLEVIROID | CbVd 1 (coleus blumei 1) | 4 | |
| CbVd 2 (coleus blumei 2) | 2 | ||
| CbVd 3 (coleus blumei 3) | 9 | ||
| AVSUNVIROIDAE | AVSUNVIROID | ASBVd (avocado sunblotch) | 33 |
| PELAMOVIROID | PLMVd (peach latent mosaic) | 98 | |
| CChMVd (chrysanthemum chlorotic mottle) | 13 | ||
| Unclassified | BSVd (Burdock stunt) | ||
| Unclassified | OFYVd (Oilpalm fatal yellowing) | ||
| Unclassified | PpMMVd (Pigeonpea mosaic mottle) | ||
| Unclassified | NGSVd (Nicotiana glutinosa stunt) | ||
| Possible viroid | CGB (Citrus Gummy Bark), possibly is CVd-II ? | ||
| Possible viroid | ELVd (Eggplant latent viroid), Avsunviroidae | Possible viroid | OOSVd (Oil palm orange spotting), Pospirviroidae |
In addition to the above classification, variants of viroids exist, and a letter is typically appended to signify the variant.
- HSVd-c (cucumber pale fruit disease)
- HSVd-g (grapevine)
The prefixes m (for "mild") and s (for "severe") are often used to designate strains, as in m–PSTVd and s–PSTVd.
The long-distance movement of PSTVd have a linguistic basis. If leaves are numbered, starting with leaf number 1, and the leaves ascending in number towards the apex leaf, then if for example, leaf 3 is innoculated with PSTVd, then only leaves above 3, and the root show symptoms of infection (that is, leaves number 1 and 2 are symptom free). Thus a grammar can easily be devised to reflect this b. We shall assume that left-to-right and right-to-left symmetries are equivalent in the following grammar.
Mover Shape Grammar
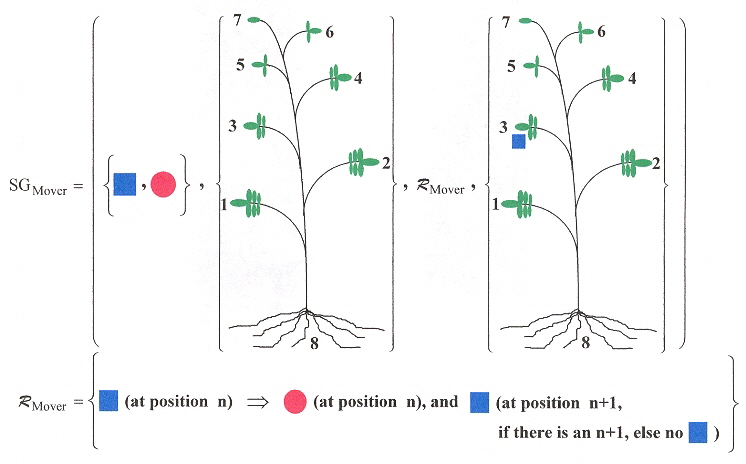
|
Example, Part B
|
Example, part C
The red markers show that viroids spread upward from the infected leaf, to the top of the plant, then infect the roots, and do not infect the leaves between the root and the originally infected leaf. |
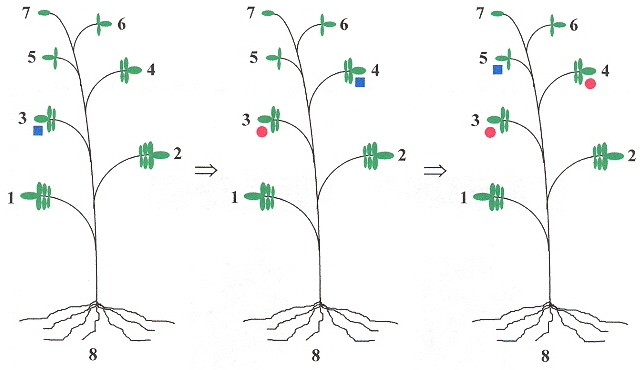
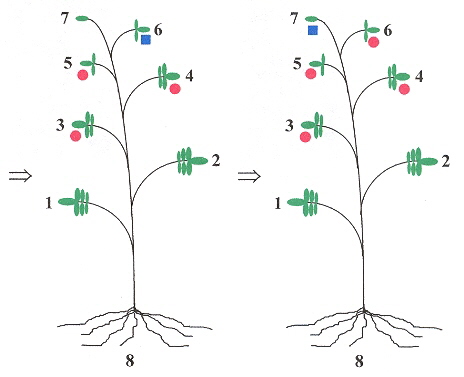
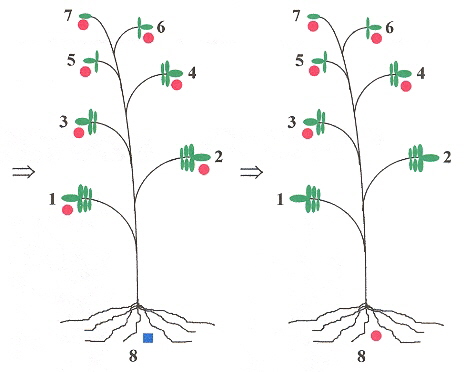
Viroids may be living fossils and have originated in the precellular RNA world c.
a The delta agent has characteristics found only in viroids and some viral satellite RNAs, suggesting that the delta agent should be classified as a viroid or a satellite RNA, and it is found enclosed in the hepatitis B virus capsid that infects human beings ("Viroids and Satellites: Molecular Parasites at the Frontier of Life", by K. Maramorosch, CRC Press, 1991, pp. 162, 163
b "Viroids", Editors: A. Hadidi, R. Flores, J. W. Randles, J. S. Semancik, CSIRO Publishing, Australia, 2003, p. 51
c "Viroids", Editors: A. Hadidi, R. Flores, J. W. Randles, J. S. Semancik, CSIRO Publishing, Australia, 2003, p. 71
"Viroids and Satellites: Molecular Parasites at the Frontier of Life", by K. Maramorosch, CRC Press, 1991, p. 3
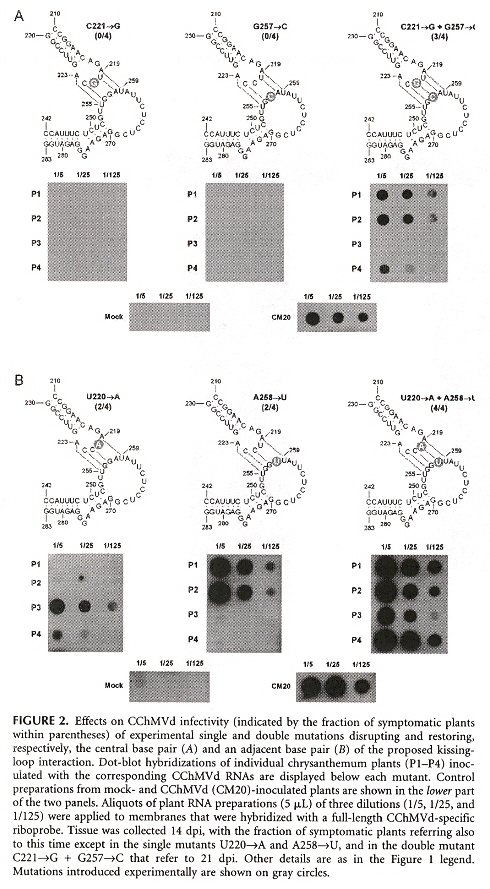
Viroids are divided into two families, Pospiviroidae (replicate in the nucleus), and Avsunviroidae (replicate in the chloroplast). Most viroids have a rod-like RNA secondary structure, but CChMVd (Chrysanthemum chlorotic mottle viroid) and PLMVd (Peach latent mosaic viroid) both support a highly branched RNA secondary conformation. These viroid RNAs probably have a pseudoknot between two hairpin loops as well. The CChMVd RNA secondary structure along with some of the experimental mutations in the RNA seconary structures, are below. The paper describes a number of other RNA mutations.
CChMVd RNA Secondary Structure
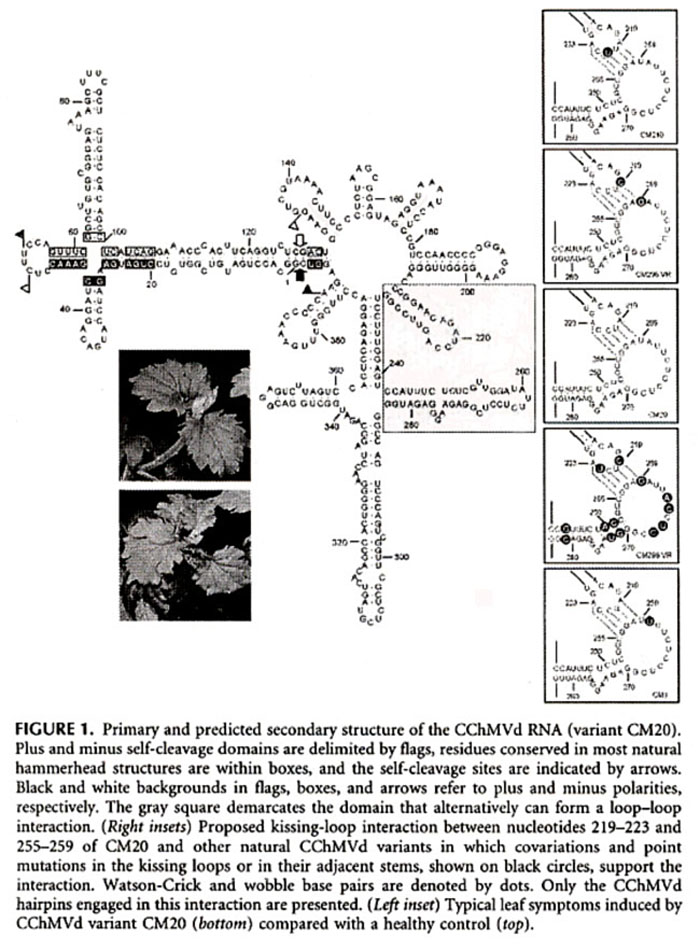
Some CChMVd RNA Mutation Secondary Structures
This paper is focused upon Posttranscriptional gene silencing (PTGS). The discovery that Peach Latent Mosaic Viroid (PLMVd), which belongs to the Avsunviroidae family (known to induce PTGS), can be used as an assay of gene silencing. One thing to result from this study is a clear explanation of what is meant by RNA sense or polarity as this assay distinguishes RNA polarity. RNA polarity is clear in the figure below.
|
Anti-sense RNA
|
RNA sense or polarity
|